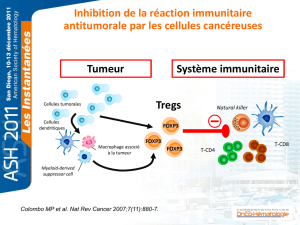
Thèse : Lymphocytes T régulateurs et GVH
Telechargé par
يوميات صيدلانية pharmacist diaries

Université Paris-Est
Ecole Doctorale Sciences de la Vie et de la Santé
Thèse de doctorat
Biologie cellulaire et moléculaire
Spécialité Immunologie
Modulation de l’effet des lymphocytes T régulateurs par la
voie TNFα/TNFR2 : une nouvelle immunothérapie en
allogreffe de cellules souches hématopoïétiques
Mathieu Leclerc
21/06/2017
Jury :
Pr. Olivier HERMINE Rapporteur
Pr. Philippe BOUSSO Rapporteur
Pr. Frédéric BARON Examinateur
Pr. Philippe SAAS Examinateur
Pr. Sébastien MAURY Co-directeur de thèse
Pr. José COHEN Directeur de thèse

1
Remerciements
Un immense merci à José. Merci pour la confiance que tu m’as accordée. Outre tes
précieux conseils tout au long de ces 3 années que j’ai eu le plaisir de passer dans
ton laboratoire, j’ai particulièrement apprécié tes grandes qualités humaines et ton
éternel optimisme! J’espère pouvoir poursuivre notre collaboration à l’avenir en
essayant notamment de transposer les résultats de ce travail, ainsi que d’autres à
venir, en pratique clinique.
Merci Sébastien pour ta confiance et ton soutien. Merci d’avoir cru en moi et de
m’avoir offert, avec José, la possibilité d’effectuer ce travail passionnant. C’est un
privilège de pouvoir travailler à tes côtés.
My Azizam, merci pour ton aide dans ce travail, qui est autant le tien que le mien.
Pendant 3 ans, on a bien bossé, mais, il faut le dire, on s’est aussi bien marré et on a
bien bouffé! Merci aussi d’avoir été le lien social le plus puissant dans ce laboratoire
et d’avoir mis une bonne ambiance au quotidien. Je te souhaite une bonne
continuation pour la suite de ta carrière de chercheur qui sera, je n’en doute pas,
brillante.
Don Kikito de la Vega, un grand merci pour ton aide et tes conseils. Ta curiosité
intellectuelle, ta générosité et ta rigueur scientifique nous ont été d’une aide
précieuse (même si tu partais parfois un peu trop en mode « fly f...er »!).
Merci bien évidemment à tout le reste de l’équipe 21 (non bis!) : Caroline, Claire,
Salim, Jérémy, Ismaïl, Marie, Cynthia, Rayane et nos « petits » étudiants qui ont
travaillé sur ce projet (Lola, Yeni, Chloé).
Un grand merci à Benoît Salomon et son équipe à la Pitié (Charlotte Pouchy, Bruno
Zaragoza) pour leur contribution à ce travail.

2
Merci à l’équipe du service de radiothérapie du Pr. Belkacemi et notamment à Jean-
Pierre Mège, Etienne Gaidier et Claude Dominique. Merci d’avoir pu vous rendre
disponible à des horaires pas toujours réjouissants...
Merci à Frédéric Charlotte d’avoir pris le temps d’analyser les (nombreuses!) coupes
histologiques réalisées au cours de ce travail.
Merci à Aurélie et Adeline, les reines du tri cellulaire.
Merci à l’équipe de l’EFPA et particulièrement à Rachid Souktani.
Merci à Marie Tamagne, Cécile Lecointe et Cindy Le Bret pour votre immense
gentillesse, votre bonne humeur si appréciable et votre volonté constante de nous
aider et de faire avancer le projet. Vous avez vraiment le cœur sur la main!
Merci à mes parents pour leur soutien indéfectible. Sans votre aide et vos
encouragements, rien de tout cela n’aurait été possible.
Enfin, last but definitely not least, le plus grand des mercis ira à mon « gang des
meufs » qui m’a supporté et encouragé tout au long de ce travail. Françounette,
Juliette et Louise, je vous aime de tout mon cœur et vous valez bien plus, fort
heureusement, que tous les Treg du monde! Et comme à cinq c’est encore mieux,
little baby is coming soon…
Je dédicace cette thèse à ma fille Louise, qui est née pendant cette parenthèse au
laboratoire et qui a déjà bien grandi et est une ravissante petite fille. Dédicace
spéciale également à Lou Reed, David Bowie et Leonard Cohen qui nous ont quittés
au cours des années consacrées à ce travail et m’ont beaucoup accompagné
pendant la réalisation des manips! Heureusement, Iggy and Neil are still here to rock
the place!
A Laurette...

3
Résumé en français
Les lymphocytes T régulateurs (Treg) jouent un rôle majeur dans la modulation de
l’alloréactivité après allogreffe de cellules souches hématopoïétiques et permettent
notamment de contrôler la réaction du greffon contre l’hôte (GVH) dans des modèles
expérimentaux. Le potentiel thérapeutique des Treg est donc très important dans ce
domaine, mais aussi dans l’auto-immunité ou en cancérologie. Cependant, de
nombreuses barrières rendent difficile l’élaboration de stratégies thérapeutiques
reposant sur le transfert adoptif de Treg chez l’homme et une meilleure
compréhension des facteurs et mécanismes contrôlant la prolifération et les
capacités suppressives de ces cellules permettrait de les cibler directement et si
possible spécifiquement in vivo.
Dans ce travail, après avoir élaboré un nouveau système d’évaluation clinique de la
GVH chez la souris et démontré sa simplicité, sa reproductibilité et sa performance,
nous avons pu montrer que l’action suppressive des Treg dans la GVH dépendait de
l’interaction entre le TNFα produit par les lymphocytes T conventionnels (Tconv) du
donneur et le récepteur TNFR2 exprimé par les Treg. En effet, en bloquant cette
interaction de 3 façons différentes, à savoir par un anticorps monoclonal bloquant
anti-TNFR2, ou en utilisant soit des Treg n’exprimant pas TNFR2 soit des Tconv ne
produisant pas de TNFα, nous avons à chaque fois montré que l’effet protecteur des
Treg était aboli en l’absence du signal TNF. Le récepteur TNFR2 étant exprimé
préférentiellement par les Treg par rapport aux Tconv, nos résultats ouvrent la voie
au ciblage des Treg in vivo via TNFR2, soit pour activer ce récepteur par un agoniste
et donc stimuler les Treg afin de contrôler la GVH, soit à l’inverse pour bloquer l’axe
TNFα/TNFR2 par un antagoniste et ainsi inhiber les Treg, ce qui permettrait alors de
lever un frein à l’alloréactivité dans les situations où l’on cherche à la stimuler pour
renforcer l’effet anti-tumoral, comme par exemple dans le cas des rechutes post-
allogreffe.

4
Résumé en anglais
Regulatory T cells (Tregs) are key players involved in the modulation of alloreactivity
after hematopoietic stem cell transplantation. Indeed, Tregs can prevent graft-versus-
host disease (GVHD) in experimental models. Therefore, the therapeutic potential of
these cells in GVHD is substantial, as it is in other fields like auto-immunity or
oncology. However, many obstacles still make the application of cellular therapy
strategies based on the adoptive transfer of Tregs in humans quite complicated. A
better understanding of factors and mechanisms that control the proliferation and
suppressive capacities of Tregs could allow for a direct and specific targeting of
these cells in vivo.
In this work, after designing a new clinical grading system for murine GVHD and
demonstrating its ease of use, reproducibility and performance, we have shown that
the suppressive action of Tregs in GVHD depends on the interaction between TNFα
produced by donor conventional T cells (Tconvs) and TNFR2 expressed by Tregs.
Using 3 different ways to block this interaction, i.e. with an anti-TNFR2 blocking
monoclonal antibody, or Tregs that do not express TNFR2 or donor Tconvs that
cannot produce TNFα, we were able to show in each situation that blocking TNF
signaling resulted in a loss of protection by Tregs. TNFR2 being highly expressed by
Tregs as compared with Tconvs, our results pave the way for in vivo targeting of
Tregs through TNFR2, either to activate this receptor with an agonist and therefore
stimulate Tregs to control GVHD, or to block the TNFα/TNFR2 axis with an
antagonist and in this case inhibit Tregs, which could boost alloreactivity, as
expected in some particular settings like post-transplant relapse.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226
227
228
229
230
231
232
233
234
235
236
237
238
239
240
241
242
243
244
245
246
247
248
249
250
251
252
253
254
255
256
257
258
259
260
261
262
263
264
265
266
267
268
269
270
271
272
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226
227
228
229
230
231
232
233
234
235
236
237
238
239
240
241
242
243
244
245
246
247
248
249
250
251
252
253
254
255
256
257
258
259
260
261
262
263
264
265
266
267
268
269
270
271
272
1
/
272
100%