Journal Identification = PNV Article Identification = 0435 Date: December 19, 2013 Time: 4:29 pm
Synthèse
Geriatr Psychol Neuropsychiatr Vieil 2013 ; 11 (4) : 379-87
Système neuromusculaire et vieillissement :
involutions et implications
Neuromuscular system and aging: involutions
and implications
Thierry Paillard
Laboratoire Activité physique
performance et santé, UPRES EA 4445,
Université de Pau & Pays de l’Adour,
Département Staps de Tarbes, France
<thierry[email protected]>
Tir ´
es `
a part :
T. Paillard
Résumé. Chez l’homme vieillissant, le nombre de cellules musculaires et d’unités motrices
décline. Les unités motrices restantes perdent de leur fonctionnalité (diminution de la fré-
quence de décharge, plus grande fluctuation de cette décharge), particulièrement celles
qui sont composées de fibres de type II. Le renouvellement des protéines intracellulaires
régresse, ce qui crée une balance négative entre les pertes protéiques journalières et les
capacités à les renouveler. L’activité de la protéine kinase (Akt) qui stimule la synthèse
de protéines de régulation (mTOR, p70S6, IGFBP-5) diminue tandis que les facteurs de
signalisation de dégradation protéique (NF-kappa B) sont activés. En outre, le processus
d’activation et de prolifération des cellules satellites est altéré et la production en hormones
anabolisantes et facteurs locaux est diminuée. Le renforcement musculaire peut contrecar-
rer ces effets délétères du vieillissement en générant une hypertrophie musculaire grâce à
une augmentation réactive de la production en hormones anabolisantes. Il peut également
engendrer une stabilisation du phénotype musculaire i.e. des différentes isoformes des
chaînes lourdes de myosine (MHC) qui évoluent sous l’effet de l’avancée en âge.
Mots clés : sarcopénie, cellule musculaire, cellule satellite, renforcement musculaire, exer-
cice aérobie
Abstract. In aged human, the number of muscle fibers and motor units decreases. The
remaining motor units lose their functionality (decrease of the discharge frequency, greater
fluctuation of the discharge) particularly those which contain type II fibers. The renewal of
intracellular proteins declines which creates a negative balance between the daily protein
losses and the capacities to renew them. The activity of the protein kinase (Akt) that stimu-
lates the synthesis of regulation proteins (mTOR, p70S6, IGFBP-5) declines whereas the
factors of degradation of proteins (NF-kappa B) are activated. Besides, the process of acti-
vation and proliferation of satellite cells is affected and the production of anabolic hormones
and local factors is decreased. After a strength training program, muscle hypertrophy is
linked to the protein synthesis at the level of myosin heavy chain (MHC) isoforms in older
subjects. However, the transcription of the genes that code the MHC-I (slow form) increases
and the transcription of the genes that code the MHC-II (fast form) decreases. Thus, the
transition of the phenotype towards a slower form cannot be inverted by strength training
during the advanced in age. Moreover, strength training enables to decrease the proportion
of fibers containing MHC of hybrid form in the process of evolution. Hence, strength training
can engender a stabilization of the muscular phenotype i.e. different isoforms of MHC. In
addition, strength training counteracts the noxious effects mentioned above by generating
muscular hypertrophy thanks to a reactive increase in the production of anabolic hormones.
A program of aerobic training can induce an increase in the synthesis of ARN messengers
coding isoforms related to the oxidative metabolism (MHC-I and to a lesser extent MHC-IIa)
while the transcribed for the type MHC-IIx decrease.
Key words: sarcopenia, muscle cells, satellite cells, strength training, aerobic exercise
Indépendamment des facteurs pathologiques, environ-
nementaux et/ou comportementaux (e.g. dénutrition,
sédentarité), la composition corporelle se modifie sous
l’effet de l’avancée en âge [1, 2]. L’atrophie musculaire
ou sarcopénie s’avère inéluctable tandis que les tissus
graisseux (adipose) et conjonctifs (fibrose) augmentent
[3].
La sarcopénie altère la fonction motrice [4] et affecte
en conséquence la qualité de vie (e.g. locomotion) et
l’indépendance physique (activités domestiques) à partir
d’un certain stade d’évolution [5]. Les effets de l’exercice
physique permettant de limiter l’ampleur du processus
sarcopénique et ses conséquences fonctionnelles sont
maintenant bien connus des scientifiques et des praticiens,
doi:10.1684/pnv.2013.0435
Pour citer cet article : Paillard T. Système neuromusculaire et vieillissement : involutions et implications. Geriatr Psychol Neuropsychiatr Vieil 2013;
11(4) :379-87 doi:10.1684/pnv.2013.0435 379

Journal Identification = PNV Article Identification = 0435 Date: December 19, 2013 Time: 4:29 pm
T. Paillard
à la fois sur les plans théorique et pratique, mais la connais-
sance de ce processus et de ses procédures préventives
évolue régulièrement.
C’est la raison pour laquelle, l’objectif de cette courte
revue est de proposer un état des lieux sur les connaissan-
ces liées à l’évolution du tissu musculaire sous l’effet du
processus sarcopénique, ainsi qu’à l’involution des capa-
cités fonctionnelles des sujets vieillissants et aux effets
mécanistiques préventifs de l’exercice physique.
Le tissu musculaire
Les fibres musculaires
Le terme sarcopénie provient du grec sarx pour «chair »
et penia pour «manque »et caractérise globalement une
perte de masse et de force musculaires. Au fil des années,
la sarcopénie traduit une dégradation des protéines muscu-
laires sous l’effet d’une altération de la protéosynthèse [6].
Elle atteint d’abord les muscles du haut du corps i.e. ceux
qui sont les moins sollicités dans la vie de tous les jours
et affecte particulièrement (et de fac¸on plus importante) le
membre inférieur à partir de 50 ans [7]. Les fibres muscu-
laires représentent 70 % du volume du quadriceps chez les
sujets de 20 ans, alors qu’elles ne représentent que 50 %
chez les sujets de 80 ans [8]. La valeur de la masse mus-
culaire resterait stable jusqu’à 50 ans, puis régresserait à
partir de cet âge pour atteindre un déficit de 30 % entre
50 et 80 ans, soit approximativement une baisse de masse
musculaire de 1 % par an [9].
Le nombre et la taille des fibres musculaires (extra-
fusales) diminuent [8]. Le nombre et la taille des myofibrilles
par fibre décroissent et le nombre de protéines contrac-
tiles (sarcomères) total par myofibrille régresse [6]. L’un des
facteurs déterminants dans la décroissance de la section
musculaire est la diminution du nombre de motoneurones-
␣(fibres myélinisées) dans les racines antérieures de la
moelle épinière (processus neuropathique) à partir d’une
soixantaine d’années. Il se produit alors une dénerva-
tion de certaines fibres [10], ce qui engendre la présence
de fibres musculaires orphelines. Une adaptation physio-
logique survient pour permettre à une majorité de ces
dernières d’être ré-innervées (les autres disparaissent) par
les motoneurones qui subsistent. Ainsi, le vieillissement
induit une augmentation du nombre de fibres par unité
motrice. En revanche, le nombre d’unités motrices décline.
Cette déclinaison apparaît liée à une diminution du fac-
teur neurotrophique ciliaire (ciliary neurotrophic factor ou
CNTF) [11]. En outre, des conclusions d’études électromyo-
graphiques soulignent une réduction du nombre d’unités
motrices fonctionnelles [8], une diminution de leur fré-
quence de décharge [12, 13] ainsi qu’une plus grande
fluctuation de cette décharge [12] chez le sujet âgé. Ceci
explique la difficulté croissante qu’éprouve le sujet âgé à
réaliser des mouvements contrôlés, fins ou précis. Ce phé-
nomène est d’autant plus marqué que la perte du nombre
d’unités motrices apparaît prononcée dans les muscles dis-
taux [14]. Avec l’avancée en âge, les individus pratiquent
de moins en moins d’activités intenses et stimulent donc
probablement moins leurs unités motrices composées de
fibres II. Ceci expliquerait en partie pourquoi, d’une part,
les motoneurones de gros calibres disparaissent dans de
plus grandes proportions que les motoneurones de petits
calibres et, d’autre part, les fibres de type II sont davantage
affectées que les fibres de type I par la dégénérescence de
leurs motoneurones. Dès lors, les fibres de type II orphe-
lines qui sont ré-innervées, le sont majoritairement par des
motoneurones de petits calibres. La nature de la commande
nerveuse influence de fac¸on déterminante les propriétés
biochimiques et mécaniques des fibres musculaires. Les
fibres de type II connectées à un motoneurone de petit
calibre se transforment progressivement sur les plans his-
tologiques et fonctionnels en fibres de type I. L’évolution
histologique musculaire évolue au fil des années et tend
vers une typologie uniforme proche de celle des fibres de
type I [15]. Globalement, le vieillissement s’accompagne
d’une diminution de la surface de section et du nombre de
fibres II [16]. Les fibres I peuvent en revanche conserver la
même surface de section et augmenter en nombre [16, 17].
Même si l’évolution typologique des fibres musculaires
n’est pas encore définitivement élucidée, on sait, cepen-
dant, que dans tous les cas, le rapport nombre de fibres
II/nombre de fibres I diminue au cours de l’avancée en âge.
Le rapport est de 1,1 à l’âge de 30 ans, puis il diminue pour
se situer aux alentours de 0,7 à 80 ans [15]. En conclusion,
la perte et l’atrophie des fibres musculaires concerneraient
principalement les fibres de type II [1, 6, 16, 18].
Sur le plan de l’architecture musculaire, le vieillis-
sement s’accompagne d’une diminution de l’angle de
pennation formé par les fibres musculaires (trajet délimité
par leurs insertions sur les aponévroses), ce qui accroît la
composante longitudinale du vecteur force et compense
partiellement la perte de force liée à l’atrophie musculaire
[19]. Cependant, la diminution du nombre de sarcomères
en parallèle (atrophie des fibres) et en série (longueur des
fibres) réduit inéluctablement la force exprimée par fibre
musculaire [6]. Le tissu tendineux subit des changements
quantitatifs et qualitatifs, lesquels affectent les propriétés
mécaniques du tendon dans la relation tension-longueur et
contribuent à détériorer la force musculaire intrinsèque du
complexe myotendineux [6].
380 Geriatr Psychol Neuropsychiatr Vieil, vol. 11, n ◦4, décembre 2013

Journal Identification = PNV Article Identification = 0435 Date: December 19, 2013 Time: 4:29 pm
Système neuromusculaire et vieillissement
Les protéines contractiles du sarcomère subissent des
involutions particulièrement au niveau du filament de myo-
sine [20]. En effet, la vitesse de synthèse protéique de la
méromyosine lourde (HMM ou heavy meromyosin), partie
constitutive des chaînes lourdes de myosine (MHC ou myo-
sin heavy chain), est ralentie avec l’avancée en âge [21]. Le
contenu relatif des formes rapides de MHC diminuant, la
proportion des formes lentes de myosine augmente. Les
isoformes des MHC d’une fibre musculaire sont uniques
chez les sujets jeunes, alors qu’elles sont de type hybride
(MHC I et MHC IIa ou MHC IIa et MHC IIx) chez les sujets
âgés [21]. La coexistence de différentes isoformes de MHC
au sein d’une même cellule musculaire chez le sujet âgé
pourrait traduire un changement du tissu musculaire sous
l’effet de l’âge. Ceci expliquerait en partie le processus de
dénervation sélectif des fibres de type II et/ou les modifi-
cations de la jonction neuromusculaire. La réduction de la
proportion des isoformes de la myosine à activité ATPasique
élevée et l’altération du couplage excitation contraction
expliqueraient une baisse sélective des fibres à contraction
rapide. De ce fait, la cinétique de la contraction musculaire
diminue [22].
La fonction mitochondriale est affectée et la mitochon-
driogenèse s’altère avec l’avancée en âge [1]. Au niveau
de la mitochondrie, on enregistre une augmentation du
nombre de lésions par délétion de fragments de l’acide
désoxyribonucléique (ADN) mitochondrial, ce qui provoque-
rait de nombreuses anomalies des complexes de la chaîne
respiratoire [9, 18]. Les effets délétères des radicaux libres
seraient à l’origine des lésions de l’ADN mitochondrial [1].
Ces mutations affecteraient le fonctionnement de la chaîne
de transport des électrons et engendreraient, par consé-
quent, la baisse de la phosphorylation oxydative [9, 18].
Par ailleurs, l’altération mitochondriale est susceptible de
constituer un signal inducteur de l’apoptose [1, 23].
Le nombre de capillaires par fibre (ratio capillaires/fibre)
régresse [24]. L’altération de la densité capillaire conjuguée
à la réduction du nombre de mitochondries induit une dimi-
nution de la capacité d’extraction de l’oxygène [25].
Le volume du réticulum sarcoplasmique diminue et sa
capacité de séquestration du calcium (Ca2+) décroît parti-
culièrement au niveau des fibres de type II [26]. En outre, sa
capacité de pompage et de relargage du Ca2+ se dégrade.
Ceci explique en grande partie pourquoi les temps de
contraction et de demi-relaxation musculaires s’élèvent au
fil du temps [26].
Au niveau cellulaire, le renouvellement des protéines
intracellulaires s’altère (le pool protéique musculaire dimi-
nue), ce qui crée une balance négative entre les pertes
protéiques journalières (100 g/jour) et les capacités à les
renouveler. L’activité de la protéine kinase (Akt) qui sti-
mule la synthèse de protéines de régulation (mTOR, p70S6,
IGFBP-5) régresse tandis que les facteurs de signalisation
de la dégradation protéique (NF-kappa B) sont activés au
cours du vieillissement [27]. L’activité de la myostatine qui
est une protéine impliquée dans l’inhibition de l’activation
des cellules satellites est augmentée [6]. Globalement, les
facteurs susceptibles de favoriser la synthèse protéique
sont atténués, tandis que les facteurs responsables de la
protéolyse sont activés.
L’étiologie de la sarcopénie demeure multifactorielle
et systémique. Un abaissement de la sécrétion en hor-
mones anabolisantes et autres facteurs de croissance telles
que la testostérone, les androgènes, les œstrogènes, la
GH (growth hormone) et l’IGF-1 (insulin-like growth factor-
type 1) ainsi qu’une élévation de l’activité de la myostatine
limitent la protéosynthèse musculaire [6]. Un état de malnu-
trition (anorexie liée à l’âge et déficience en vitamine D) et
un manque d’activité physique limitent également les fac-
teurs activateurs de la synthèse protéique. En revanche, la
présence de plus en plus fréquente au cours de l’avancée
en âge de cytokines [interleukine-1 (IL-1), interleukine-6
(IL-6) et du facteur de nécrose tumorale (TNF)], qui sont
des médiateurs sécrétés par les macrophages lors d’états
inflammatoires, témoigne de leur rôle actif dans le pro-
cessus du vieillissement [28]. L’élévation de ces facteurs
immunologiques reflète l’augmentation de la protéolyse
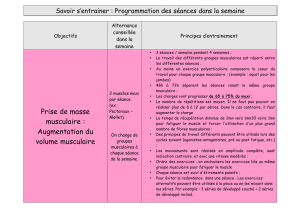
avec l’avancée en âge [6] (figure 1).
Au niveau des structures sensorielles du muscle, les
fuseaux neuromusculaires subissent également des chan-
gements morphologiques et fonctionnels. Le diamètre des
fuseaux neuromusculaires diminue [29], certaines fibres
intrafusales, essentiellement les fibres à chaînes nucléaires,
disparaissent tandis que l’épaisseur de la capsule du
fuseau augmente [30, 31]. En conséquence, la sensibi-
lité des fuseaux neuromusculaires régresse, ce qui affecte
l’efficacité du réflexe myotatique [32].
Sur le plan génétique, il existerait des gènes spécifiques
qui marquent des différences entre plusieurs individus au
niveau du phénotype musculaire. La nature de celui-ci
aggrave ou atténue le processus sarcopénique.
Les cellules satellites
Ce sont des myoblastes quiescents situés en périphé-
rie des myocytes dont la fonction est de fusionner avec
eux pour réparer leur lésion (un nombre de noyaux supé-
rieur à la normale dans une cellule musculaire montre
qu’une cellule satellite a fusionné avec elle) ou de les
remplacer (formation des nouvelles fibres) lorsqu’ils se
nécrosent. Le vieillissement s’accompagnerait intrinsèque-
ment (en dehors des facteurs environnementaux) d’une
Geriatr Psychol Neuropsychiatr Vieil, vol. 11, n ◦4, décembre 2013 381

Journal Identification = PNV Article Identification = 0435 Date: December 19, 2013 Time: 4:29 pm
T. Paillard
Processus neuropathique
et apoptose Protéosynthèse musculaire Protéolyse musculaire
Dénervation des myocytes (+++)
Ré-innervation des myocytes (+)
Pertes motoneurales Facteurs
hormonaux
Facteurs
environnementaux
Testostérone ↓
Androgènes ↓
Oestrogènes ↓
GH et IGF1 ↓
Myostatine ↑
Statut oxydant/
anti-oxydant
Facteurs
immunologiques
Nombre d’unités motrices ↓
Nombre de fibres ↓
Taille des fibres ↓
Régénération des fibres ↓
Masse musculaire ↓ (sarcopénie)
(+++) : importante ; (+) : insuffisante (ne compense pas les dénervations)
Activité physique ↓
Apports protéiques ↓
Radicaux libres ↑
Anti-oxydants ↓
IL-1β ↑
IL-6 ↑
TNF-α ↑
Figure 1. Représentation schématique du processus sarcopénique.
Figure 1. Schematic representation of the process of sarcopenia.
régression du pouvoir d’action des myosatellites [33]. Bien
que les cellules satellites des sujets âgés puissent main-
tenir leur capacité à répondre aux facteurs mitogéniques
(e.g. facteurs de croissance des fibroblastes basiques ou
bFGF et protéine S100B), elles demeurent néanmoins dans
l’incapacité de les sécréter [33]. Ceci contribue à dimi-
nuer le nombre et la taille des fibres musculaires par
déficit de régénération. Cette altération du pouvoir de
division et de différenciation semble également liée au
nombre de cycles dégénérescence/régénération auxquels
les cellules satellites ont participé. Au cours de la vie d’un
homme, le nombre de division des myosatellites semble
déterminé génétiquement. Le pool de myosatellites dimi-
nuerait donc avec l’avancée en âge. Le nombre de cellules
satellites par fibre musculaire serait effectivement plus
faible chez les sujets âgés comparativement aux sujets
jeunes, mais le nombre de noyaux cellulaires par fibre serait
plus élevé [34]. Ces auteurs précisent que le nombre de
myosatellites comparé au nombre total de noyaux (myo-
satallites/noyaux + myosatellites) serait plus faible chez les
sujets âgés comparativement aux sujets jeunes. En outre,
le vieillissement se caractériserait par une altération du pro-
cessus d’activation et de prolifération des cellules satellites
[1, 6, 35]. Ceci pourrait expliquer la perte de masse mus-
culaire liée au vieillissement et l’altération de la capacité de
régénération.
Toutefois, une autre hypothèse est avancée pour
expliquer l’altération croissante du pouvoir d’action des
myosatellites. En fait, le défaut d’activation des cellules
satellites pourrait être lié à une modification de leur environ-
nement, laquelle serait engendrée par des diminutions de
la production hormonale, des facteurs locaux et de l’activité
musculaire [9], mais également par un changement du sta-
tut oxydant et anti-oxydant de l’organisme.
Les capacités fonctionnelles
La force musculaire
La force développée par un muscle est proportionnelle
à sa surface de section. Cette dernière diminue et contri-
bue à la réduction inéluctable de la force maximale d’un
sujet au fil des années. La masse et la force musculaires
déclinent parallèlement de 30 à 50 % entre 30 et 80 ans
[36]. Dès lors, il convient de s’intéresser au rapport force
musculaire/masse musculaire. Bien que certains auteurs
aient montré qu’il ne décline pas [37], Frontera, et al. [38] ont
plus récemment observé qu’il pouvait régresser lorsque les
structures protéiques contractiles subissent des involutions
(surtout chez les hommes). Ce phénomène semble asso-
cié à une atrophie sélective des fibres de type II [39]. Ces
fibres sont en effet celles qui développent le plus de force.
Par ailleurs, pour une surface de section donnée d’une fibre
musculaire atrophiée, la diminution de la force serait davan-
tage liée à une perte du nombre de ponts acto-myosine qu’à
une réduction de force de chaque pont d’union [6].
La capacité de production de force est davantage
affectée pour les mouvements rapides que pour les mou-
vements lents [40]. Le pourcentage de perte de puissance
382 Geriatr Psychol Neuropsychiatr Vieil, vol. 11, n ◦4, décembre 2013

Journal Identification = PNV Article Identification = 0435 Date: December 19, 2013 Time: 4:29 pm
Système neuromusculaire et vieillissement
musculaire (-3à4%/an) serait même deux fois plus impor-
tant que celui de perte de force maximale (-1à2%/an) [41].
Ce phénomène semble lié à un ralentissement de la ciné-
tique contractile [42]. Les sujets âgés entraînés développent
une puissance musculaire supérieure aux sédentaires du
même âge. Toutefois, l’involution de la puissance rappor-
tée au volume musculaire, est régulière et parallèle pour
des populations actives et sédentaires [43].
Par ailleurs, chaque contraction musculaire s’accom-
pagne d’une coactivation des muscles agonistes et anta-
gonistes. Le rôle des muscles antagonistes est d’assurer
une meilleure stabilité articulaire et une meilleure action
des muscles agonistes. Le vieillissement du système ner-
veux central peut également être tenu responsable de la
dégradation de la coordination entre les muscles agonistes
et antagonistes [44]. Ce phénomène peut réduire la force
musculaire et la précision du mouvement chez le sujet
âgé. Chez un sportif, l’entraînement diminue l’activation des
antagonistes, ce qui élève le niveau de force musculaire
résultant des agonistes et facilite l’apprentissage de coordi-
nations spécifiques (mouvements complexes). Cependant,
le vieillissement se caractérise par un phénomène (inverse)
de désadaptation. Ainsi, le degré de la coactivation des
muscles antagonistes augmente avec l’âge (perturbation de
l’inhibition réciproque) et réduit les capacités de production
de force musculaire et de contrôle du mouvement.
La réponse évoquée par le réflexe H régresse égale-
ment sous le poids des années. Dans la mesure où le
réflexe H emprunte la même voie nerveuse que le réflexe
d’étirement, bien qu’il ne transite pas par les fuseaux neu-
romusculaires, cette observation indique que l’excitabilité
de la boucle spinale diminue sous l’effet de l’âge [45]. La
régression de ces réflexes altère les capacités des sujets
âgés à réagir efficacement et rapidement à des perturba-
tions physiologiques ou mécaniques pouvant mettre en péril
leur équilibre postural. En effet, l’altération de la fonction
sensorielle myotatique altère la proprioception et dégrade
en conséquence la fonction d’équilibration ce qui contribue
à augmenter le risque de chute chez la personne âgée.
Globalement, les involutions fonctionnelles du système
neuromusculaire sont susceptibles d’affecter la qualité de
vie des personnes âgées dans le cadre de leurs activités
physiques domestiques quotidiennes telles que les mon-
tées d’escaliers ou les levées de chaise. Par ailleurs, on a
rattaché l’augmentation de la fréquence des chutes avec
l’âge, à une diminution de la force et de la masse mus-
culaires du membre inférieur. Néanmoins, une régression
modérée de la force musculaire aggrave peu le risque de
chute tant qu’un seuil critique n’est pas atteint. En revanche,
une fois ce seuil franchi, le risque augmente notablement
[4].
La fatigabilité musculaire
Certains auteurs prétendent que les sujets âgés sont
moins résistants à la fatigue que les sujets jeunes [46, 47],
d’autres présentent des résultats inverses [48, 49]. D’autres
encore suggèrent que le régime de contraction de la tâche
fatigante influencerait les résultats [50]. Théoriquement,
la persévération des fibres de type I devrait entretenir la
qualité d’endurance musculaire chez le sujet vieillissant.
Effectivement, le ralentissement des propriétés contractiles
et l’évolution progressive d’un métabolisme préférentielle-
ment oxydatif engendrerait une économie énergétique et
une réduction d’accumulation de métabolites inhibitoires
[51]. Le métabolisme devenant davantage oxydatif et moins
glycolytique au fil des années favoriserait une plus grande
résistance à la fatigue chez les sujets vieillissants. Cepen-
dant, les effets du vieillissement pourraient également
induire de profondes modifications structurelles et fonction-
nelles au niveau des mitochondries (cf. paragraphe les fibres
musculaires). Ces modifications ne sont pas favorables à
une plus grande résistance à la fatigue (i.e. une améliora-
tion des capacités d’endurance) chez les sujets âgés. En fait,
la santé, le style de vie des sujets (sujets actifs vs sujets
non actifs) ainsi que la nature des protocoles expérimentaux
(e.g. actions musculaires isométriques ou dynamiques, dif-
férentes vitesses de contraction) impactent probablement
fortement les résultats dans l’étude de la fatigabilité au
cours de l’avancée en âge.
Les effets de l’exercice
Le renforcement musculaire
L’entraînabilité des sujets âgés demeure puisqu’un pro-
gramme de renforcement musculaire de douze semaines
comprenant 3 séances hebdomadaires composées de
mouvements d’extension du genou et de flexion du coude
(3 séries de 8 répétitions à 80 % de 1 RM -répétition maxi-
male), a induit des résultats édifiants chez des sujets âgés
de 60 à 72 ans [52]. En effet, à l’issue du programme, la
force d’extension a augmenté en moyenne de 107 % et
la force de flexion de 227 %. Selon le groupe musculaire
considéré (ou entraîné), l’amplitude des adaptations peut
varier sensiblement.
L’adaptabilité musculaire de la femme semble être sen-
siblement identique à celle de l’homme à l’égard des
exercices de renforcement musculaire pour une population
de personnes âgées. Lexell, et al. [53] ont en effet observé
des augmentations de force sans différence significative
entre des hommes et des femmes âgés de 70 à 77 ans
à l’issue d’un programme de 11 semaines. Les forces de
Geriatr Psychol Neuropsychiatr Vieil, vol. 11, n ◦4, décembre 2013 383
 6
7
8
9
6
7
8
9
1
/
9
100%