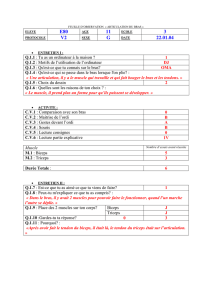
Relations Force-Vitesse des composants musculo-tendineux

!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
Université de Nantes, UFR STAPS
Relations Force-Vitesse des composants
musculo-tendineux du triceps sural
Mémoire de recherche, Master 2 spécialité « Expertise, Performance,
Intervention »
2011/2012
Présenté par HAURAIX Hugo
Directeurs de recherche : Dorel Sylvain, Nordez Antoine

Remerciements :
Je tiens tout d’abord à remercier, Antoine Nordez et Sylvain Dorel, mes deux directeurs de
mémoire qui m’ont permis de mener à bien ce travail. Je n’oublie pas l’ensemble des
enseignants chercheurs de l’UFR STAPS qui m’ont aussi aidé dans cette tâche.
Merci à Kévin, Mathieu, Lilian, Hugo, Killian, Tristan, Martin, Agathe et Marie Cécile pour
cette ambiance et convivialité au sein du bureau des doctorants.
Enfin, je remercie l’ensemble des sujets qui ont participé à cette étude.

!
0!
1 Introduction ........................................................................................................................ 1
2 Revue de littérature ........................................................................................................... 2
2.1 Relation force-vitesse ............................................................................................................... 2
2.2 Étude du comportement de l’ensemble muscle-tendon à partir de l’échographie en
temps réel ............................................................................................................................................ 3
2.3 Impact de l’interaction muscle-tendon .................................................................................. 6
2.4 Relation force-vitesse et utilisation de l’échographie ............................................................ 8
2.5 Différences entre les muscles du triceps sural ....................................................................... 9
2.6 Apport de l’échographie ultra-rapide .................................................................................. 10
3 But de l’étude et hypothèses ............................................................................................ 10
4 Matériels et méthode ........................................................................................................ 11
4.1 Sujets ....................................................................................................................................... 11
4.2 Matériels ................................................................................................................................. 11
4.2.1 Ergomètre .......................................................................................................................... 11
4.2.2 Observation des fascicules musculaires et des jonctions myo-tendineuses ...................... 12
4.3 Protocole ................................................................................................................................. 13
4.4 Traitement des données ......................................................................................................... 13
4.4.1 Calcul des données anthropométriques ............................................................................. 13
4.4.2 Calcul des forces ............................................................................................................... 13
4.4.3 Analyse des images échographiques ................................................................................. 14
4.4.4 Interpolation des données .................................................................................................. 14
4.4.5 Calcul des paramètres ....................................................................................................... 15
4.5 Analyse statistique .................................................................................................................. 16
5 Résultats ............................................................................................................................ 17
5.1 Répétabilité des mesures ....................................................................................................... 17
5.2 Comportement des différents composants du triceps sural ............................................... 17
5.2.1 Données globales .............................................................................................................. 18
5.2.2 Gastrocnémiens médial ..................................................................................................... 18
5.2.3 Gastrocnémien latéral ....................................................................................................... 19
5.2.4 Variation des différents éléments de l’ensemble muscle-tendon ...................................... 20
5.3 Relations Force-Vitesse des différents composants du triceps sural ................................. 21
5.4 Vitesses de raccourcissement maximales des fascicules musculaires ................................ 23
6 Discussion .......................................................................................................................... 23
6.1 Comportements des différents composants musculo-articulaire ....................................... 23
6.2 Relations Force-Vitesse .......................................................................................................... 25
6.3 Raccourcissement des fascicules musculaires ...................................................................... 26
6.3.1 Différence des vitesses de raccourcissement maximales des fascicules musculaires entre
le gastrocnémien médial et latéral ................................................................................................. 26
6.3.2 Variabilité inter-individuelle des valeurs de raccourcissement maximale des fascicules
musculaires .................................................................................................................................... 27
6.4 Considérations méthodologiques .......................................................................................... 28
6.4.1 Cinématique du mouvement ............................................................................................. 28
6.4.2 Calcul de la force .............................................................................................................. 28
7 Conclusion et perspectives .............................................................................................. 30
8 Bibliographie .................................................................................................................... 31
9 Annexes ............................................................................................................................. 37

!
1!
1 Introduction
Dans le cadre de mon Master 2 recherche « Expertise, Performance, Intervention »,
j’ai effectué mon stage au sein du laboratoire « Motricité, Interactions, Performance » (EA
4334) à l’UFR STAPS de Nantes. Ce laboratoire pluri-disciplinaire possède deux axes
thématiques majeurs, ce mémoire s’inscrit plus particulièrement dans l’axe « Adaptations de
la fonction neuromusculaire ».
A l’heure actuelle, la notion de relation force-vitesse est très utilisée dans le monde de
l’entraînement sportif pour caractériser le profil d’un sportif par des qualités de « vitesse » ou
bien de « force ». La notion de puissance maximale est, elle aussi, très utilisée dans
l’entraînement, elle est égale au produit de la force et de la vitesse. Cette information de
puissance maximale est corrélée à la performance lors de sprints courts ou de squat (Morin et
al., 2002). Il est donc intéressant de regarder l’évolution de cette relation après un protocole
d’entraînement longitudinal (Cormie et al., 2010) ou bien après un protocole de fatigue (Jones
et al., 2010). Il existe aussi plusieurs autres domaines d’application où la relation force-vitesse
donne une information sur le sujet. Le domaine de la réhabilitation et celui de la gérontologie
y trouvent un intérêt particulier pour voir dans le premier cas, l’état de capacité du sujet à
reprendre une activité normale, et dans l’autre, observer l’effet de l’âge sur les différents
paramètres musculo-articulaire (Raj et al., 2010).
Même si la relation force-vitesse globale à de nombreuses fois été étudiées, les
relations force-vitesse des sous-systèmes du complexe musculo-tendineux on quant à elles
rarement été rapportées dans la littérature. Ce mémoire s’inscrira donc dans ce cadre, afin de
mieux comprendre l’adaptation du comportement des éléments de l’ensemble muscle-tendon
avec l’augmentation de la vitesse de raccourcissement du système musclo-tendineux.

!
2!
2 Revue de littérature
2.1 Relation force-vitesse
La vitesse d’exécution d’un mouvement va dépendre de la force qui est exercée sur le
système (Goubel et Lensel-Corbeil, 2003). Plus la force appliquée au système est importante,
plus la vitesse du mouvement sera faible (figure 1). Les travaux précurseurs au sujet de la
relation force-vitesse ont été effectués sur muscle ou bien fibres isolées (Fenn et Marsh,
1935). Hill, en 1938, a proposé une équation permettant de modéliser la relation force-vitesse
dans le but d’estimer par extrapolation la vitesse maximale de raccourcissement sous charge
nulle (figure 1). Toutefois, le comportement « in vivo » ne peut être interprété de la même
façon que le comportement obtenu sur muscle isolé (Ahn et al., 2006).
Figure 1: Relation entre la force normalisée et la vitesse de raccourcissement d’un
fascicule musculaire avec une extrapolation de cette relation par l’équation proposée par
Hill afin d’estimer la vitesse maximale de raccourcissement (Ahn et al., 2006).
« In vivo », Différents protocoles existent afin d’obtenir une relation couple-vitesse, et
ce à partir de mouvements poly-articulaires, comme lors d’une tâche de pédalage
(Vandewalle et al., 1987) ou lors d’un mouvement de squat (Limonta et al. 2010). Il est aussi
possible d’extraire une relation couple-vitesse à partir, cette fois-ci, de mouvements mono-
articulaires effectués sur ergomètre isocinétique (Ingemann et Halkjaer, 1979 , Reeves et
Narici, 2003).
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
1
/
52
100%