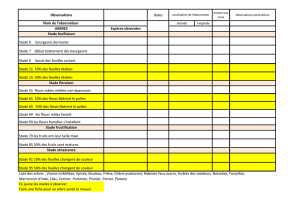
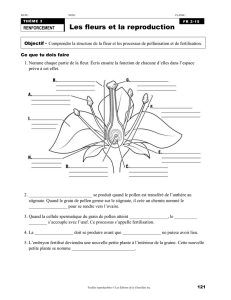
Les angiospermes

Organisation du vivant Végétal
Les angiospermes
I] Introduction
Les angiospermes alternent phase haploïde (gamétophyte) et diploïde (sporophyte). L'apparition d'organes sexuels (fleurs) est
une évolution, issue de la modification de feuilles pour la protection. Depuis le XVIII° siècle, la théorie de la métamorphose
florale a été posée : la fleur n'est qu'une tige modifiée. Actuellement, cette théorie est totalement vérifiée : nombreuses
similitudes au point vue moléculaire et anatomique (vascularisation, forme...)
II] Morphogenèse florale
Généralement, un phytomère végétatif se transforme en phytomère d'inflorescence, qui portera plusieurs fleurs. Dans de rares
cas, le phytomère végétatif devient directement floral.
Transition florale → Acquisition de la réceptivité aux facteurs inductif → Morphogenèse florale
1) La transition florale
Au stade végétatif, les méristèmes caulinaires sont incapables de recevoir les
facteurs environnementaux inductifs de la floraison.
Les facteurs physiologiques de cette transition florales sont : l'âge (varie selon
les espèces), la température (vernalisation), la lumière (photopériode), mais
aussi des signaux endogènes (hormones florigènes).
Photopériode : un seuil critique de réception de lumière (durée
essentiellement) permet la différenciation florale, mais c'est en fait la durée de
l'obscurité qui est perçue.
•Héméropériodiques : jours longs
•Nyctiopériodique : jours courts
Le flash lumineux entraîne l'absence de floraison chez les plantes de nuit. La réception de la photopériode se fait au niveau de
la feuille, la lumière déclenchant l'utilisation de l'amidon stocké pour augmenter la concentration en saccharose qui entraînera à
son tour la production de cytokinines.
Florigène : la greffe d'une feuille induite sur une plante non induite entraîne la floraison. Cependant, il n'a jamais été possible
d'isoler une telle substance, laissant donc le florigène au stade d'hypothèse.
Vernalisation : le traitement au froid permet la floraison.
2) Évocation florale
Se fait au niveau du méristème. Il subit
trois phases de développement : phase
juvénile, phase adulte (peut faire une
fleur quand les conditions sont réunies),
phase reproductive.
Le méristème passe d'une phase de
croissance indéterminé à une phase de
croissance déterminée par perte de
l'anneau primaire. Élargissement du
sommet du méristème, augmentation du
volume des nucléoles. La zone centrale
est indifférenciée en division, une zone
périphérique.

II] Diversité de l'organisation florale
1) Morphogenèse florale
Périanthe (partie stériles) : pétale et sépales chez les hétérochlamydée. S'il
manque l'un des deux, monochlamydée.
Parties fertiles : étamines (androcées) et carpelles (gynécées).
Le pédoncule florale porte la fleur, la bractée florale constitue la face
ventrale, et axe inflorescent la face dorsale.
L'apparition de ces différentes pièces est sous contrôle génétique ( fonctions
A,B,C) :
•A : sépale
•A+B : Pétale
•B+C : Etamine
•C : Carpelle
L'inactivation de ces gènes donne des fleurs composées de feuille (tige non différenciée.
2) Les modes de répartition des sexes
Le plus fréquemment on rencontre des fleurs monoclines (hermaphrodites). Les fleurs dicline sont soit mâle, soit femelle. Les
plantes monoïques possèdent les deux sexes, alors que les dioïques ont les sexes sur des pieds différents.
3) Morphologie des fleurs
Les cycliques : Les pièces sont organisées en verticille de l'extérieur vers l'intérieur → sépales, pétales, étamines, carpelles
Acyclique : les pièces organisées en spirale
Hémicycle : sépale et pétales sur verticille, carpelle et étamines sur spirale
Chez les monocotylées chaque cycle contiendra 3 pièces (trimères). Chez les eudicotylées on aura des tétramère ou pentamère.
Si le nombre de pièces par cycle est identique, on parle de cycles isomères, opposés au cycles hétéromères. Le plus
communément, on rencontre des fleurs tétracyclique ou pentacyclique (4 ou 5 cycles). Les symétries peuvent êtres biradiales,
radial (actinomophique), bilatérale (zygomorphe), et asymétriques.
Diagramme : on représente les tissus chlorophylliens en hachure. On commence par orienter (bractée floral, axe inflorescent).
Attention aux verticille, et leur potentiel décalage.
Formule florale : f (+) = 5S + 5P + 5E1 + E2 + (3C) (les parenthèses indiquent que les carpelles sont liées).
•Fleur zygomorphe (bilatérale) : F = 5S + [(5P) + 4E] + (2C)
•Pisum sativum : F = (5S) + 3P1 + (2P) +[5E1 + 4E] + (2C)
Périanthe : ensemble des pièces qui protègent les organes.
Ces ensembles vont être caractéristiques des différentes familles.
a) Inflorescences monopodique
La disposition des fleurs sur la tige d'une plante est souvent caractéristique d'une famille.
1er cas de figure d'Inflorescence monopodique : Croissance indéterminé, production latérale de fleurs. Les fleurs les plus
âgées sont en bas (acropètes, opposé à acrofuge).
α) Fleurs pédonculées
•Alternée
◦Fleurs étagées → Grappe
◦Fleurs dans même plan → Corymbe (Longueur du pédoncule = âge de la fleur)
•Verticillé → Ombelle

β) Fleurs sessiles
•Fleurs périanthées :
◦Axe allongé : Epi
◦Axe allongé et charnu : Spadice
◦Axe ramassé et charnu : Capitule
•Fleurs apérianthées :
◦Fleur monocline (Hermaphrodite) : Epillet (Graminées)
◦Fleur dicline :
▪Axe allongé : Chaton
▪Axe ramassé et charnu : Sycone (Figuier)
γ) Sympodique
Le bourgeon au lieu de s'allonger, va donner une fleur.
•Fleurs pédonculées :
◦Dichasiales (bipares) : Cyme bipare (Centaurium erythraea)
◦Monochasiales (unipares) :
▪Alternée : Cyme hélicoïdale (Hemerocallis fulva)
▪Non alternée : Cyme scorpioïde (Myosotis arvensis)
δ) Composées
On peut avoir des système pédonculées à étage : Grappe d'épis, ombelle d'ombelles, et Corymbe de Capitules
III] Androcée
1) Morphologie d'une étamine
L'étamine est l'organe de reproduction mâle constitué d'un filet (fin pédicelle) surmonté par une anthère (constituée de 2 loges
polliniques, donc 4 sac polliniques). La jonction entre les deux s'appelle le connectif. La microsporogenèse se passe dans ces
sacs polliniques.
Il existe divers formes de ces étamines en rapport avec la position du connectif et celle des anthères :
a) Basifixe ou innée
b) Médifixe
c) Dorsifixe
d) Apicifixe
e) A thèques décalées
f) Monothèque (médifixe)
Les pièces fertiles (étamines) peuvent être : Libre ou soudées par les filets (Gamostémonie) ou les anthères (Synanthérée)
2) Ontogenèse staminale
Primordium staminale formera les anthères. Il existe 3 assises :
L1 → épiderme + stomium
L2 → cellules archésporiales
L3 → tissus conducteurs et connectif
L'archéspore va se diviser pour former la cellule pariétale, des cellules de
l'endothélium, des assises transitoires, tapis staminal (en contact avec le
tissu sporogène, on suppose que tapis va servir à l'alimentation de ce
tissu). La cellule sporogène donnera le tissu sporogène (2n)
Toutes ces formation donneront l'anthère.

a) Maturation staminale
Elle va démarrer dans le tissu sporogène par une méiose. Chacune des cellules de ce tapis va subir une méiose, pour former
une structure en tétrade (formée de 4 cellules haploïdes) avec une paroi en callose. Le début de la maturation s'opère dès que
les cellules du tapis sont capable de produire la protosporopollènine. L'hydrolyse de la callose (grâce à une glucanase) entraine
une libération des microspores.
Différenciation d'une paroi pollinique, le sporoderme : constitué de 2 paroi (Exine + Intine). L'exine étant elle même
constitué d'une ectexine contenant la sporopollènine et d'une endoxine riche en polysaccharide.
Cette différenciation aura un rôle dans la
dissémination (ex : petit et lisse → plus facilement
balayé par le vent, grand avec des poils → rôle
adhésif sur les insectes/sol)
Formation du gamète mâle : gonflement de la
cellule, et excentrement du noyau. La cellule
générative (plaqué contre la paroi), et la cellule
végétative (grosse cellule). La cellule générative va
se coller à l'intine et s'internaliser. Le grain de
pollen est composé de deux cellules (70% des
angiospermes). Les 30% restants sont composés de
grain de pollen tricellulé, formées par deux cellules
génératives. Une seconde mitose se fait après
formation du pollen.
Le pollen est largué par déhiscence des antères (rupture du sac pollinique).
Certains types cellulaires de l'anthère subiront une destruction séquentielles.
La déhiscence est due à une différenciation d'une assise mécanique, puis à
une dégénérescence des tissus les plus internes, entraînant la fusion de deux
sacs polliniques.
IV] Le gynécée
a) Différences de réalisation d'un système ovarien
Les carpelles peuvent être libres (apocarpie) ou partiellement fusionnés (syncarpie). La soudure entre les carpelles
(concrescence) peut se faire uniquement au niveau de l'ovaire, mais peut se prolonger jusqu'au style.
Le placenta relie les ovules au système nourricier.
Il y a plusieurs types de placentation :
•Marginale : repliement des bords
•Pariétale : soudure des parois les unes
avec les autres (ovules au niveau des
soudures)
•Axile
•Centrale
Chez les Brassicacées, placentation pariétale, avec
fausse paroi centrale.

b) Rapport du gynécée avec les autres pièces florales
Si l'ovaire est placé au dessus du cycle d'insertion des
pièces florales (hypogyne) : on parle de gynécée supère. Si
l'ovule est entouré par les autres pièces (périgyne) : on
parle de infère. Si les pièces sont au dessus de l'ovaire
(épigyne) : on parle d'infère.
c) L'ovule
Le sac embryonnaire contient les cellules fertiles. Il est protégé par le nucelle, lui
même protégé par les téguments interne (secundine) et externe (primine). Le hile
est la zone de raccord entre les téguments et , la chalaze est la base du nucelle qui
correspond à la zone ou le tissu conducteur diverge pour irriguer les téguments. Le
mycropyle est une petite ouverture en position apicale (point d'entrée du tube
pollinique). Le funicule par du placenta pour irriguent l'ovule. Le raphé est la zone
de soudure entre ovule et funicule.
On peut avoir plusieurs courbures :
•Anatrope : comme ici, avec retournement total
•Orthotropes : droit
•Campylotrope : courbure à 90° entre hile chalaze et
micropyle.
d) Ontogenèse de l'ovule et macrosporogenèse
La cellule archésporiale sous l'épiderme du nucelle va subir une première division, la
cellule la plus externe (pariétale) va avorter. La cellule restante (sporogène) va subir
une méiose, les trois cellules les plus externes vont également avorter. La plus interne
(mégaspore) donnera le sac embryonnaire (mégagamétogenèse). La mégaspore va
subir 3 mitoses successives sans cellularisation (séparation de l’ADN), donc donnant
une cellule à 8 noyaux. Cette cellule se cellularise en 7 cellules : 3 antipodes, 2
synergides, une oopshère (zygote principal) et une cellule centrale diploïde (albumen
triploïde après fécondation). Il faut donc deux fécondation pour féconder l'oosphère.
V] Pollinisation et double fécondation
a) Autogame
20% des angiospermes sont autogame strictes. Très utile lorsque l'on travaille sur des plantes à intérêt agronomiques (pas de
perte des génotypes), et permet une colonisation rapide depuis un seul individus. Cependant, ce type de fécondation conduit à
une impasse évolutive. Chez les violettes par exemple, la fleur est fertile avant son ouverture (cléistogames).
b) Allogame
Avantage évolutif avec la vigueur hybride (hétérosis). Avantage dans les environnements changeants. Permet des fécondation
« à distances », avec donc nécessité d'évolution du pollen pour la dispersion. Les espèces dioïques sont forcément allogames.
Les fleurs peuvent présenter des incompatibilités morphologiques (long style) ou avoir besoin de vecteurs de pollinisation :
•Insectes (entomogamie) : pollen lourd et ornementé, avec morphologie particulière.
•Vent (anémogamie) : nécessite un pollen léger, peu ornementé, pollinisation de masse
•Eau (hydrogamie)
•Oiseau (ornithogamie) : nécessite du pollen lourd et très ornementé pour s'accrocher, et morphologie de la fleur pour
déposer le pollen sur l'animal
•Les mammifères : homme, chauve souris
Chez les allogames hermaphrodites, il y a de nombreux phénomène d'auto-incompatibilité pour éviter l'autogamie : auto-
incompatibilité au niveau génétique (l'oosphère empêche la progression du tube pollinique), ou niveau sporophytique
(reconnaissance de la décoration du pollen empêchant la germination).
 6
7
6
7
1
/
7
100%