III - Appareil reproducteur chez les phanérogames

LSV1 – Semestre 2 – Biologie de la reproduction et du développement végétal – appareil reproducteur chez les phanérogames - 1
BIOLOGIE DE LA REPRODUCTION ET DU
DEVELOPPEMENT VEGETAL
APPAREIL REPRODUCTEUR CHEZ LES
PHANEROGAMES
Nicolas Pauly, pas cours le 15 avril, cours repoussé au 22 avril.
INTRODUCTION
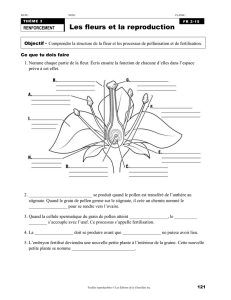
QU’EST CE QU’UNE FLEUR ?
Après les parties végétatives, on s’intéressera aux parties reproductives d’une plante : à un moment donné se
met en place, par un mécanisme de transition florale, une phase reproductive. A l’issue de cette phase
reproductive, apparaissent des fleurs produisant des gamètes mâles et femelles, des hormones, etc. La
rencontre des gamètes est particulière dans la mesure où elle est double : deux gamètes sont fécondés, puis il
y a mise en place d’un embryon, à l’origine du retour à une phase végétative.
L’étude de la germination est très pratiquée via les recherches et méthodes permettant la compréhension et
l’amélioration des plantes ayant un intérêt humain. C’est en cela que la transition florale est un point clé : on
arrête la production de la phase végétative et de sa biomasse, et on met en place des organes reproducteurs
ainsi que des graines. Il est dans l’intérêt de l’Homme d’être en mesure de comprendre les mécanismes
permettant de retarder la transition florale, permettant ainsi la production d’une plus grande biomasse. Ce
partie
végétative
partie
reproductrice
embryon
germination

LSV1 – Semestre 2 – Biologie de la reproduction et du développement végétal – appareil reproducteur chez les phanérogames - 2
sont ces points clés qui sont les plus étudiés par les botanistes. Ces observations permettent néanmoins d’aller
au-delà de l’étude intéressée et de comprendre les mécanismes moléculaires en jeu.
Les angiospermes représentent approximativement 170 000 espèces actuellement. La structure de
reproduction est la fleur. Celle-ci va se développer à partir d’un bourgeon terminal. Elle consiste en un rameau
raccourci portant des feuilles modifiées d’un point de vue fonctionnel et architectural, permettant d’assurer la
fonction de reproduction.
• Une fleur consiste tout d’abord en des pièces stériles :
o les pétales ont une fonction d’attraction grâce à des pigments, des molécules odoriférantes,
et ce plutôt lorsque la fleur est au stade bouton
o des sépales, ayant un rôle de protection
• Mais aussi des pièces fertiles :
o Les étamines
o Le pistil ou gynécée
CE QU’EN PENSE GOETHE…
D’une fleur à l’autre, des caractéristiques différent : nombre de pièces florales, couleur, disposition, etc. Ces
clés de base ont permis la mise en place de la classification traditionnelle. L’origine de la définition de la fleur
remonte à Goethe, en 1790, au sein de son œuvre « le traité de la métamorphose », précurseur en la matière.
Dans les années 1990, on validait ses théories et notions grâce aux nouvelles technologies, qui stipulaient
notamment que les parties florales présentent malgré tout de nombreuses ressemblances avec les parties
végétatives, et qu’une fleur n’est rien d’autre qu’une tige modifiée. Les observations étaient les suivantes :
• Chez les angiospermes primitives, observation des pièces florales disposées en spirale de la même
façon que les feuilles sur la tige.
• La vascularisation des sépales et pétales est identique à celle d’une tige feuillée.
• Les organes reproducteurs, en particulier sur les étamines, sont pétaloïdes, induisant que les étamines
sont aussi des feuilles modifiées.
• Les structures sont en apparence très différentes, mais finalement, le modèle initial pour leur mise en
place semble être commun avec, au cours de la partie finale de la construction, une orientation vers
les parties végétatives ou reproductrices.
Toutes les angiospermes produisent des graines ayant des intérêts au sein de l’agro-industrie. Beaucoup de
travaux (par exemple les croisements de variétés), dans notre région, ont eu et ont toujours un facteur
économique important de par un passé tourné vers l’industrie florale.

LSV1 – Semestre 2 – Biologie de la reproduction et du développement végétal – appareil reproducteur chez les phanérogames - 3
MI SE A FLEUR ET MOR PHOGENESE FLORALE
Les méristèmes caulinaires vont subir
d’importants changements leur permettant une
transformation vers des méristèmes floraux.
Au sein des plantes, et par analogie avec la
segmentation dans le monde animal, on
retrouvera trois types de phytomères
possédant chacun leurs spécificités :
• Métamères végétatifs
• Métamères inflorescentiels : concerne
les structures ramifiées et les inflorescences de
la plante
• Métamères floraux : on les retrouvera
sur les inflorescences formées
Il peut exister que le passage d’un métamère végétatif à un métamère floral se fasse sans passage par le
métamère inflorescentiel, comme par exemple chez la tulipe. Ce passage peut être divisé en trois phases :
• Transition ou induction florale : concerne l’acquisition de la compétence à être réceptif aux facteurs
inductifs de la floraison
• Evocation florale : séquence des évènements qui interviennent dans le méristème caulinaire en
réponse à un processus inductif
• Morphogénèse florale : au sens strict du terme, c’est la formation des pièces florales.
INDUCTION FLORALE
Initialement, les méristèmes sont totalement
incompétents au sein de la plante, c'est-à-dire
incapables de répondre aux facteurs
physiologiques et environnementaux qui
induisent la mise en fleur, et ce car ces
méristèmes ne sont pas armés pour répondre
à tous les changements endogènes ou
exogènes de la floraison. En d’autres termes,
même si les conditions physiologiques ou
environnementales sont optimales, on n’aura
jamais de transition florale. Quels sont ces
facteurs importants ?
ÂGE
Tant que la plante n’a pas atteint un stade végétatif suffisamment avancé, elle sera incompétente à la
transition florale. C’est un facteur interne.

LSV1 – Semestre 2 – Biologie de la reproduction et du développement végétal – appareil reproducteur chez les phanérogames - 4
• Chez la tomate, la transition florale n’aura jamais lieu avant l’apparition sur le plan de 13 phytomères
successifs.
• Chez le seigle, il faudra non pas des entrenœuds mais 7 feuilles. Pour les arbres en particulier, ce
temps de maturation peut être important.
• Arabidopsis thaliana est une plante modèle regroupant des caractéristiques de travail intéressantes
physiologiquement et génétiquement. Son génome est le mieux séquencé. On peut obtenir facilement
obtenir des mutants, en particulier floraux. Cette plante nécessite une rosette de quatre feuilles.
PHOTOPERIODE
La lumière sera un facteur permettant
cette transition. La photopériode
correspond à l’alternance entre phases
sombres et phases éclairées, sur 24
heures. En plus de la période de
luminosité, on doit introduire la notion de
quantité de lumière mais aussi de spectre
ou qualité de la lumière. Il existe pour
chacune des plantes un seuil critique
permettant ou non l’induction florale et
les autres phases de la mise à fleur. Il
existe deux grands types de
photopériode :
• Les plantes de jour long, ou plantes héméropériodiques, telles qu’Arabidopsis thaliana, la moutarde
ou la jusquiame.
• Les plantes de jour court ou nyctipériodiques telles que Nicotiana tabacum, Xanthium (lampourde),
Pharbitis (volubilis).
SIGNAUX HORMONAUX
En effectuant chez une plante une transition jour court-
jour long, on observe un enchaînement d’évènements
dès perception du jour long au niveau foliaire :
1. Perception du jour long.
2. Les réserves amidacées sont mobilisées en
saccharose.
3. Le saccharose est véhiculé au travers de la sève
élaborée par le phloème.
4. Il est transporté en particulier vers le méristème
apical racinaire
5. La perception de ce saccharose en cet endroit
est un signal induisant la synthèse de cytokinine, une hormone végétale.
6. L’hormone sera véhiculée dans les parties aériennes au niveau des feuilles, mais surtout au niveau du
méristème apical caulinaire, le tout avec un temps de latence de 9h.
7. Le méristème apical caulinaire est baigné de cytokinine permettant l’induction florale sans retour.

LSV1 – Semestre 2 – Biologie de la reproduction et du développement végétal – appareil reproducteur chez les phanérogames - 5
LA VERNALISATION
La vernalisation est une transformation interne opérée par le froid qui confère à certaines plantes l’aptitude à
fleurir. Il n’y aura pas de modification morphologique.
Chez le blé, il existe des blés d’hiver, et des blés de printemps. Ce sont les sélectionneurs, qui, sur plusieurs
dizaines d’années, ont permis l’obtention de ces deux variétés. Via la vernalisation, le blé d’hiver est plus
précoce, et est d’un meilleur rendement. On le sème à l’automne, et il va passer l’hiver à l’état de jeune
plantule, fleurir au printemps, et épier au début de l’été. On le récolte début juillet. S’il ne subit pas le froid, il
ne fleurit pas. Le blé de printemps, quant à lui, est semé au printemps, il va fleurir en été, passe l’hiver et fera
des épis au printemps.
Via la vernalisation, il existera trois classes de plantes :
• Plantes indifférentes (certaines céréales, tabac)
• Plantes préférentes : la vernalisation n’est pas indispensable mais hâte la floraison
o Chez le seigle d’hiver, plus de vernalisation induit une floraison dès 7 feuilles, et moins de
vernalisation une floraison dès 25 feuilles.
• Plantes à vernalisation obligatoire : plantes bisannuelles (betteraves, choux, carottes, olivier). En cas
de vernalisation absente, il n’y a pas de floraison. Bouh.
o Chez l’olivier, celui-ci a besoin d’un abaissement thermique pour pouvoir produire des fruits :
la température doit descendre en dessous de 10°C. La localisation de l’olivier d’un point de
vue géographique est donc assez réduite : une altitude trop élevée ne conviendra pas à la
plante, étant fragile au froid, et une altitude trop basse empêchera toute floraison, les
températures n’étant pas assez basses.
La transition florale est clairement multifactorielle.
EVOCATION FLORALE
On se situera ici d’un point de vue
tissulaire. La croissance du méristème
végétatif est indéterminée tant qu’il n’a
pas subi de transition florale. Lorsque
celle-ci survient, l’aspect est
relativement différent, conduisant à la
formation d’un méristème floral à
croissance déterminée générant une
fleur terminale. On retrouvera donc lors
de l’évocation florale une activité
mitotique plus importante, qui va avoir
tendance à bomber l’aspect général du
méristème floral, sur lequel on
retrouvera en partie supérieure une
structure en éventail. Ces structures ne
sont donc que des feuilles modifiées formant les différentes pièces florales. Globalement, on peut donc dire
que lors de l’évocation florale a lieu un bouleversement au niveau cellulaire du méristème, mettant en place
méristème floral et pièces florales.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
1
/
26
100%