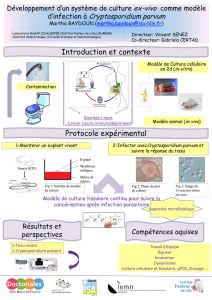
Differenciation in vitro de blastodermes de Truite (Salmo irideus

Differenciation
in
vitro de blastodermes de Truite
(Salmo irideus) depourvus de couche enveloppante
par CH. DEVILLERS, J. COLAS, et L. RICHARD1
Laboratoire d'Anatomie et d'Histologie de la Faculte des Sciences, Universite de Paris
L'ETUDE
structural
et
fonctionnelle du blastoderme de Poisson conduit
a y
distinguer deux ensembles: la couche enveloppante et les blastomeres profonds.
Si du seul point de vue quantitatif (volume total des cellules) la couche envelop-
pante est beaucoup moins importante que la masse blastomerienne, du point de
vue qualitatif elle represente
un
element essentiel dans
le
developpement
du
blastoderme depuis
le
stade indivis jusqu'a
la
fermeture
du
blastopore. C'est
en elle que resident les proprietes d'impermeabilite, de coordination des mouve-
ments cellulaires, de contractilite et d'expansibilite d'ensemble du blastodisque
qui entrent en jeu a divers stades de la premorphogenese et de la morphogenese.
Vis-a-vis de ce revetement 'actif le groupe blastomerien profond fait plutot figure
de masse inerte, ou tout au moins incoherente, incapable 'd'initiative' et dont les
possibility's morphogenetiques, en particulier les mouvements lors de la gastrula-
tion, ont besoin d'etre coordonnees, dirigees par la couche enveloppante. Mais
c'est
de cet
ensemble que derive
la
quasi-totalite
du
materiel cellulaire
qui
edifiera l'embryon;
la
couche enveloppante ne constituera que l'assise
la
plus
superficielle de l'epiderme, apres avoir fourni au materiel de la zone marginale
un appoint cellulaire, presque negligeable en quantite.
De
la
cooperation de ces deux constituants resultent
la
mise en place
et la
differenciation de l'embryon. Si nous empechons cette cooperation qu'adviendra-
t-il de la morphogenese?
Une telle epreuve peut etre tentee en debarrassant le blastoderme de sa couche
enveloppante et laissant la masse des blastomeres profonds adherente au vitellus.
Par suite de l'intervention de mouvements de reaction du syncytium vitellin et de
la pellicule perivitelline, les resultats de cette experience, souvent contradictoires,
sont difficiles a interpreter. II apparait alors plus simple de faire la meme opera-
tion sur des blastodermes separes de leur vitellus; par cette methode nous avons
obtenu quelques resultats qui valent d'etre analyses.
1
Authors' address: Laboratoire d'Anatomie
et
d'Histologie Comparees, Faculte" des Sciences,
1 Rue Victor Cousin, Paris, France.
[J.
Embryol. exp. Morph. Vol. 5, Part 3, pp.
264-273,
September 1957]

DIFFERENCIATION DE BLASTODERMES DE TRUITE 265
MATERIEL ET METHODE
Des oeufs
de
Salmo irideus sont places quelques heures dans
une
solution tres
diluee
de
rouge neutre,
ce qui
permet
de
limiter pratiquement
la
coloration
du
blastoderme
a la
seule assise cellulaire
de la
couche enveloppante. Cette colora-
tion prealable facilite beaucoup
la
conduite
de
l'operation.
La
coque
est
ensuite
ouverte
et la
couche enveloppante
est
'pelee'
a
l'anse
de
platine, operation
relativement facile
sur
l'oeuf
de
Truite; avec
un peu
d'habitude
il est
possible
d'enlever
la
couche d'une seule piece
en
n'entrainant avec elle
que peu de
blastomeres profonds. Couche enveloppante
et
masse
de
blastomeres profonds
detaches
du
vitellus, sont alors cultives
en
godets separes
a la
temperature
de
7-8°.
Le
liquide
de
culture
est le
milieu
de
White (1946) reduit
a sa
portion
minerale
et a son
glucose
et
dont
la
teneur
en Ca a ete
amenee
au
meme taux
que dans
le
liquide
de
Holtfreter:
Dextrose 0,85
g.
NaCl 0,7
g.
KC1 0,0375
g.
CaCl2 0,029 g.
MgSO4 0,0275
g.
FeNO
3
0,14 mg.
Na2HPO4 0,0145
g.
KH2PO4 0,026
g.
Eaubidistilleeq.s.
. . .
100 cm.3
Le grattage
de la
couche enveloppante peut etre effectue
aux
stades morula
a
petites cellules, blastula, tres jeune gastrula. Passe ce dernier stade
le
toit blasto-
celien devient trop mince pour qu'on puisse arracher
la
couche sans entrainer
avec elle les 3
ou 4
assises cellulaires sous-jacentes.
RESULTATS
(1) Blastomeres profonds
Sur
50
cultures effectuees
il n'a ete
obtenu
que 6
resultats analysables, tous
a
partir de jeunes gastrulas.
En
effet, les explants
de
blastomeres profonds (blasto-
meres grattes)
peu
coherents
se
dissocient souvent
et
surtout
se
necrosent plus
rapidement, semble-t-il, que ceux des blastomeres complets (c'est-a-dire pourvus
de leur couche enveloppante). Deja la survie et la differenciation des blastodermes
complets sont des phenomenes capricieux; bien souvent ces explants se necrosent
ou survivent sans se differencier, sans que nous puissions preciser les raisons d'un
tel comportement. Dans
le cas des
explants grattes
ou
nous multiplions
les
chances
de
'leser'
le
blastoderme,
il
n'est pas etonnant que
les
resultats soient
si
maigres.
Des explants grattes
de
morula
ou de
blastula
ont pu
survivre parfois dans
d'assez bonnes conditions de conservation pendant
5-7
jours, mais se presentent
seulement comme
des
amas informes sans traces
de
differentiations. Tout
au

266 DEVILLERS ET AL. — DIFFERENCIATION IN VITRO DE
plus observons-nous des tentatives de groupements cellulaires, separes du reste
de la masse par des limites plus ou moins nettes correspondant a des changements
d'orientation des cellules. Le blastocele se constitue dans les explants de morula.
Dans l'ensemble la morphologie est comparable a celle des 'hyperblastulas'
(Oppenheimer,
1936)
realisees par
les
blastodermes complets preleves tres jeunes.
Une autre constatation, valable pour tous les explants quel que soit leur stade de
prelevement, qu'ils soient ou non differencies, est que la couche enveloppante ne
se reconstitue pas. II peut en subsister des fragments qui se reconnaissent facile-
ment sur la surface de l'explant par la forme et l'alignement regulier de leurs
FIG.
1. Coupe d'un explant de blastomeres profonds pre'leve' au
stade gastrula jeune et fixe 5 jours apres. c, corde; ce, fragments
de couche enveloppante ayant subi regalement Epibolique; s, sur-
face de l'explant form6e par les blastomeres profonds.
cellules. Dans les explants gastruleens ces ilots de couche enveloppante sont
entres en activite epibolique comme le temoignent l'aplatissement et retirement
de leurs cellules (Fig. 1). Une telle activite ne se manifeste dans aucun explant
pregastruleen alors que des ceufs temoins, fixes a la meme date que ces explants,
sont en pleine epibolie.
Si nous examinons maintenant les quelques explants qui ont subi des differen-
ciations nous noterons d'abord certaines particularites de leur morphologie
generate comparee a celle d'explants complets, differencies, du meme age.

BLASTODERMES DE TRUITE 267
Tandis que l'explant complet se roule, en quelques heures, en une sphere parfaite
a surface lisse, par un mouvement du surtout a l'activite de la couche envelop-
pante (Devillers, 1951!)), l'explant gratte ne subit qu'un lent modelage par
remaniement et accroissement des contacts intercellulaires et finalement il
esquisse vaguement la forme d'une sphere a surface rugueuse dont les cellules
superficielles arrondies ne presentent aucune ordonnance reguliere et peuvent
raeme se detacher. De tels explants conservent cette forme jusqu'a la fin de leur
survie sans jamais subir l'allongement rencontre si souvent avec les blastodermes
complets.
La differenciation la plus nette, obtenue avec les jeunes gastrulas, est celle du
tissu cordal (Fig. 1), bien reconnaissable a la forme de ses hautes cellules,
empilees, ecrasees les unes contre les autres, a contenu plus clair que celui des
autres cellules. Le degre de differenciation de ce tissu atteint parfois celui de la
corde produite par des blastodermes complets mais plus generalement il est
moins pousse; l'aspect est plus embryonnaire et les limites de ces fragments de
corde sont moins bien tranchees par rapport au reste de l'explant. Du point de
vue topographique ce materiel cordal presente deux caracteristiques: d'une part
il n'a guere subi d'elongation et d'autre part il se rencontre a l'etat de noyaux
plus ou moins importants, isoles les uns des autres, soit en pleine masse de l'ex-
plant, soit en surface, c'est-a-dire avec une localisation tout a fait anormale.
Les somites sont apparus mais avec une differenciation toujours moins bonne
que celle de la corde. On peut en reconnaitre des series, assez reduites en
longueur, ou la metamerie se distingue et ou l'ordonnance des cellules est assez
caracteristique.
Les meilleurs s'alignent en courtes series ou la metamerie se manifeste assez
bien; le groupement des cellules dans chaque somite y est bien caracteristique.
Mais d'autre part, dans la masse de l'explant se rencontrent de nombreuses
formations, isolees, qui montrent une disposition radiaire des cellules autour
d'une zone centrale, pleine ou creusee d'une petite lumiere. Dans certains cas il
s'agit
incontestablement de somites, mais pour nombre d'entre eux la determina-
tion devient beaucoup plus difficile et douteuse.
En ce qui concerne la topographie nous pouvons, comme pour la corde,
observer la dispersion de ces differenciations dans toute la masse et la surface
de l'explant. D'autre part elles ne presentent pas les relations typiques avec les
formations cordales.
Un troisieme groupe de differenciations est constitue par des structures
neuroides (Fig. 2). Ce sont des masses irregulieres a noyaux arrondis, plutot
groupes a la peripherie et laissant vers le centre une zone de cytoplasme clair.
Jamais il n'a ete rencontre de formations nerveuses embryonnaires typiques,
immediatement reconnaissables.
Enfin des formations tubulaires ont ete observees a plusieurs reprises. Elles
sont en general courtes mais dans un cas, un tel tubule, bien organise*, ferme aux
deux extremitds, occupe toute la hauteur de l'explant. La lumiere centrale y est

268 DEVILLERS ET AL. — DIFFERENCIATION IN VITRO DE
bien nette, la paroi composee, sur presque toute sa longueur, d'une seule assise
cellulaire. La determination en est difficile; il pourrait
s'agir
de formations du
type lame laterale.
En surface, les fragments de couche enveloppante subsistant, se sont etires et
presentent les noyaux allonges, caracteristiques de cette structure lorsqu'elle
accomplit son mouvement epibolique, mouvement favorise ici par l'existence
d'un substratum.
n
FIG.
2. Meme explant que dans la fig. 1. n, masse neuroi'de;
som,
differenciation de type somitique.
(2) Couche enveloppante
Tres rapidement l'explant de couche enveloppante se roule en une sphere
parfaite qui subsiste sous cet etat un certain temps, puis, a un moment qui doit
correspondre, dans l'histoire de l'oeuf normal, a la mise en route de I'epibolie, la
forme commence a se modifier; l'explant s'allonge et sa surface se herisse de
plissements (Fig. 3) qui sont le temoignage d'un etalement de la couche envelop-
pante, correspondant au processus epibolique, incompletement realise ici, par
suite del'absence de substratum sur lequel les cellules puissent adherer et ramper.
Ces dernieres conservent alors une forme plus arrondie.
Les coupes ne montrent aucune differenciation dans ces explants, qui pourtant
comportent une portion de zone marginale. Peut-etre la quantite de ce materiel
est-elle trop faible ou bien les cellules constitutives de cette region ont-elles aban-
donne la couche, realisant, in vivo, le processus d'ingression observe sur l'oeuf
normal (Devillers, 1951a). II n'est pas possible d'en decider.
 6
7
8
9
10
6
7
8
9
10
1
/
10
100%