syllabi - Fédération Wallonie

COURS GENERAUX
« Moniteur sportif éducateur »
THEMATIQUE 3
Facteurs déterminants de l'activité
et de la performance
MODULE 3
La "machine humaine" à l'effort

CG2_Th3_Mod2_La « machine humaine » à l’effort Page 1 sur 62
COURS GENERAUX
« Moniteur sportif éducateur »
THEMATIQUE
3
Facteurs déterminants de l'activité et de la
performance
MODULE
3

CG2_Th3_Mod2_La « machine humaine » à l’effort Page 2 sur 62
La "machine humaine" à l'effort
Cours généraux de la formation « Moniteur Sportif Educateur »
Thématique 3 : Facteurs déterminants de l’activité et de la performance
Module 2 : La « machine humaine » à l’effort
BENOIT Nicolas, Centre d’Evaluation de la Performance Sportive (CAPS), UCL
FRANCAUX Marc, Centre d’Evaluation de la Performance Sportive (CAPS), UCL
TONDEUR Alexandra, Faculté des Sciences de la Motricité, UCL
Fédération Wallonie-Bruxelles, Administration Générale d’Aide à la Jeunesse, de la Santé et du Sport,
Direction Générale du Sport (Adeps), Service « Etudes et développements de projets ».
adeps.formationdecadres@cfwb.be
⇒ R
ESUME
:
Ce module de formation est le prolongement du module CG1_Th3_Mod1_La machine humaine. Pour
rappel, dans le cadre d’une formation dont l’objectif est de préparer de futurs moniteurs qui auront
comme vocation l’encadrement de sportifs, la compréhension de la « machine » dont l’individu va se
servir pour pratiquer son sport est indispensable.
Au premier stade de formation, la « machine humaine » a été décrite de façon très générale.
L’objectif de ce module-ci se veut plus spécifique par rapport à l’exercice physique. En effet, une
série de systèmes ont, petit-à-petit, été mis en évidence pour leur rôle essentiel lors d’un exercice
physique. Une importance qui se concrétise essentiellement par une adaptation spécifique lors de
l’exercice mais aussi, pour certains d’entre eux, par une adaptabilité plus grande lors de la répétition
de ces exercices (entraînement). Les 3 systèmes physiologiques principaux qui interviennent à
l’exercice ainsi que les mécanismes et les modifications sous-jacents seront donc décrits plus en
détails.
Au terme de cette unité de formation, le candidat « moniteur sportif éducateur » devrait donc
pouvoir comprendre et connaître les bases de fonctionnement du corps humain en activité
physique.

CG2_Th3_Mod2_La « machine humaine » à l’effort Page 3 sur 62
⇒ M
ETHODOLOGIE
:
- Exposé magistral
- Séance « questions-réponses »
⇒ S
UPPORTS DE COURS
:
- Syllabus
- Foire aux questions (FAQ)
- Présentation assistée par ordinateur (PAO)
⇒ M
ODALITES D
’
EVALUATION
:
- Questionnaire à choix multiple (QCM)
⇒ C
HARGE THEORIQUE DE TRAVAIL POUR LE CANDIDAT
:
- En présentiel :
5 heures de cours magistral
1 heure d’évaluation
- En non présentiel :
12 heures d’étude indépendante et personnelle en guise de préparation à l’évaluation

CG2_Th3_Mod2_La « machine humaine » à l’effort Page 4 sur 62
Ce module de formation apportera des réponses aux questions suivantes (liste non exhaustive):
De nombreuses sous-questions sont envisagées afin de permettre au lecteur une meilleure
compréhension des sujets abordés.
NB : Il est fortement recommandé au candidat moniteur sportif éducateur de suivre la
présentation orale de ce module de cours. De très nombreux exemples permettront
certainement au candidat moniteur sportif éducateur d’appréhender encore plus concrètement
ces notions qui, de prime abord, peuvent paraître rébarbatives !
M
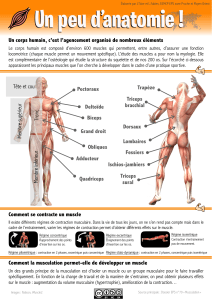
USCLE ET MOUVEMENT
Qu’est-ce qu’un muscle ? (p 8)
Quels sont les différents types de contraction musculaire ? (p 8)
Comment la contraction musculaire est-elle contrôlée ? (p 9)
Comment l’information neuronale est-elle transmise ? (p 10)
La commande motrice est-elle hiérarchisée ? (p 10)
Comment l’information neuronale est-elle transformée en contraction musculaire ? (p 11)
Qu’est-ce qu’une unité motrice ? (p 12)
Quel est le rôle des unités motrices dans le développement de la force ? (p 13)
Existe-t-il une adaptabilité du contrôle moteur ? (p 13)
Quelle est la structure microscopique du muscle ? (p 15)
Pourquoi le nom de muscle squelettique strié ? (p 16)
Quelle est la structure microscopique d’un sarcomère ? (p 16)
Quel est le mécanisme microscopique de la contraction musculaire ? (p 18)
Comment les filaments fins et épais savent-ils à quel moment ils doivent interagir et former
des ponts transversaux pour permettre au muscle de se contracter ? (p 20)
Qu’est-ce que la relation Force-Vitesse ? (p 20)
Quelle est l’utilité de la relation Force-Vitesse dans l’entraînement ? (p 20)
Qu’est-ce que la typologie musculaire … ? (p 20)
L
E SYSTÈME CARDIO
-
VASCULAIRE
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
1
/
63
100%