1/16 07/03/2016 MANDON Bastien L3 CR : BOUACHBA

SYSTÈME CARDIOVASCULAIRE – Physiologie cardio-vasculaire
1/16
07/03/2016
MANDON Bastien L3
CR : BOUACHBA Amine
Système cardio-vasculaire
Pr Stéphane DELLIAUX
16 pages
Physiologie Cardio-vasculaire
A. Introduction-Généralités
Andrea Cesalpino (1519-1603) a défini le terme de circulation. Le cœur a pour rôle de faire circuler le sang. Il
introduit également la notion de mouvement perpétuel.
La fonction du cœur est de mobiliser le sang et de permettre la circulation, ceci afin de transporter l'O2 et les
nutriments vers les tissus et les cellules, puis de collecter le CO2 et les produits du métabolisme afin de les
acheminer vers les organes épurateurs (rein, foie, poumons).
La machinerie cardiaque est un moteur bien huilé : on compte environ 60 battements par minute soit
2 à 4.109 battements tout au long de la vie.
C'est un organe vital à l'origine de morbidité et de mortalité. Il est autonome et possède une activité propre. Il
est régulé par différents systèmes comme le système neurovégétatif (SNV) ou encore différentes hormones.
L'embryogenèse du cœur est très importante car celui-ci est composé de différents tissus ayant différentes
origines.
On y retrouve :
• Essentiellement du muscle strié viscoélastique et contractile. Ces cellules musculaires striées sont
excitables et se comportent comme un syncytium, c'est à dire que ce qui se passe dans une cellule est
transmis aux cellules adjacentes.
• Du tissu fibreux, qui sert à la structure de cœur et à la séparation des différentes cavités.
• Du tissu électrogénique c'est à dire du tissu de conduction électrique ayant la capacité d'initier un
potentiel d'action.
Plan
A. Introduction-Généralités
B. La contraction cardiaque
I. L'électrogénèse cardiaque
II. Le couplage excitation-contraction
III.Le raccourcissement cardiomyocytaire
C. Le cycle cardiaque
I. Généralités
II. Le cycle diastole-systole
III.Le diagramme de Wiggers
D. Énergétique cardiaque
I. Consommation cardiaque en oxygène
II. La contraction non chargée
III.La contraction chargée

SYSTÈME CARDIOVASCULAIRE – Physiologie cardio-vasculaire
2/16
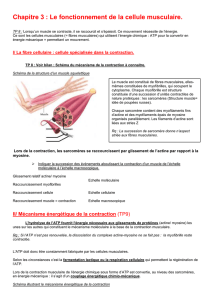
B. La contraction cardiaque
I. L'électrogenèse cardiaque
a) Le tissu nodal
Le tissu nodal, ou tissu cardionecteur, est regroupé dans des structures de deux types, des paquets ou nœuds et
des faisceaux. C'est lui qui permet la naissance de l'activité électrique du cœur.
Les cellules de ce tissu ont gardé certaines capacités provenant des cellules embryonnaires. Elles ont donc des
propriétés particulières :
• Automaticité : Capacité de remplir sa fonction en l'absence de stimulation extérieure
• Rythmicité : Sa fonction se répète de manière cyclique
• Excitabilité : Capacité de générer une activité électrique et de se dépolariser.
• Auto-excitabilité : Peut générer un phénomène électrique à type de potentiel d'action (PA) sans
stimulation extérieure
• Conductibilité : Capacité de transmettre un phénomène électrique à une autre cellule
• Contractilité : Capacité de se contracter et de générer une tension ou une force aux extrémités
On retrouve deux structures nodales : Le nœud sinusal et le nœud atrio-ventriculaire.
Au niveau des faisceaux on retrouve le faisceau de His ainsi que ses branches droite et gauche, les faisceaux
inter atrio-ventriculaires et le réseau de Purkinje.
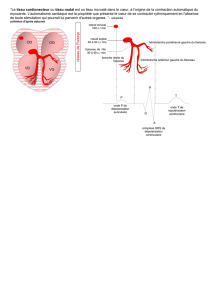
b) L'auto-excitabilité du tissu nodal
L'auto-excitabilité est la capacité
d'autogénérer un phénomène électrique de
type potentiel d'action.
Si l'on regarde la cellule sinusale (cellule
du tissu nodal), on remarque qu'entre
chaque PA le potentiel de repos est instable
et croissant. Lorsque le potentiel de repos
dépasse un certain seuil, aux alentours de -
40mV, il se génère automatiquement un
potentiel d'action.
Dans les cellules ventriculaires par contre
le potentiel de repos est stable et constant.
Il n'y a pas de tendance spontanée à la
dépolarisation, donc pas de génération
spontanée de potentiel d'action.
L'autoexcitabilité est ainsi une particularité des cellules nodales.

SYSTÈME CARDIOVASCULAIRE – Physiologie cardio-vasculaire
3/16
c) La dépolarisation diastolique lente
Lors de la diastole on remarque un phénomène particulier appelé dépolarisation diastolique lente.
C'est une dépolarisation lente des cellules
nodales causée par 3 phénomènes ioniques :
• Le courant de fond
• Le courant pacemaker
• Le courant calcique transitoire
Le courant de fond est dû à la Na+/K+ ATPase et à l'échangeur Na+/Ca2+ permettant l'entrée de sodium et la
sortie de calcium.
Dans la phase de dépolarisation diastolique lente il y a une activation de l’échangeur Na+/Ca2+ ainsi qu'une
diminution de l'activité de la Na+/K+ ATPase. On constate ainsi une augmentation de la concentration de
sodium intra-cellulaire, ainsi qu'une sortie de calcium.
Le calcium intra-cellulaire est très présent en début de diastole. Ainsi c'est sa forte concentration qui
conjointement à l'entrée de sodium va participer à la dépolarisation diastolique lente.
Le courant pacemaker est dû à des canaux ioniques HCN. Ce sont des canaux non spécifiques qui dépendent
du degré d'hyperpolarisation de la cellule.
Ils s'activent lors d'une hyper-polarisation de la membrane. Ainsi plus la cellule sera hyperpolarisée, plus ces
canaux seront actifs.
Ces canaux HCN sont perméables au sodium et au potassium. Ainsi en phase de diastole, il y aura une entrée
progressive de sodium d'autant plus que la membrane sera hyperpolarisée.
Le courant calcique transitoire est dû à un canal calcique de type T voltage dépendant. A partir d'un certain
niveau de dépolarisation ce canal est activé et perméabilise la membrane cellulaire pour le calcium, provoque
son entrée et donc une dépolarisation.
C'est la réunion de ces 3 phénomènes (courant
de fond, courant pacemaker et courant calcique
transitoire) qui permettent la dépolarisation
spontanée diastolique lente de la membrane.
Le tissu nodal a une tendance à la dépolarisation
spontanée dépendante de sa localisation au niveau
du cœur. Schématiquement plus le tissu nodal est
« haut » dans le cœur, plus celui-ci aura une
fréquence de dépolarisation rapide.

SYSTÈME CARDIOVASCULAIRE – Physiologie cardio-vasculaire
4/16
On remarque que la pente de dépolarisation du tissu nodal est différente selon la localisation.
Ainsi :
• La tendance spontanée à la dépolarisation est plus élevée au niveau sinusal, l'onde de dépolarisation
surviendra prioritairement à son niveau. C'est donc le rythme spontané du nœud sinusal qui impose le
rythme cardiaque sinusal à environ 60bpm.
• Si le nœud sinusal est détruit, le deuxième élément à pouvoir prendre le relais est celui ayant la pente de
dépolarisation la plus rapide. Ce sera donc le nœud atrio-ventriculaire qui va prendre le relais à un
rythme cardiaque jonctionnel de 30-40bpm.
• Si ce nœud est également détruit, on verra apparaître un rythme cardiaque idioventriculaire de 10-
20bpm provenant du tissu nodal ventriculaire.
Cette différence de fréquence spontanée du tissu nodal selon sa localisation cardiaque est due à une différence
phénotypique et morphologique de ce tissu au sein du cœur.
C'est l'ensemble de ces phénomènes de dépolarisation et repolarisation qui vont donner l'aspect de l'ECG avec
ses ondes P, T et le complexe QRS.
d) La conduction de l'excitation cardiaque
La dépolarisation naît normalement au niveau du nœud sinusal. Elle se propage et atteint le nœud atrio-
ventriculaire avant de passer dans les cavités ventriculaires cardiaques. Suite à la très haute conductivité du
tissu nodal dans le faisceau de His, la dépolarisation des ventricules naît à l'apex et se poursuit vers la base
des ventricules
• L'onde P est le témoin de la dépolarisation auriculaire.
• L'onde Q est le début de la dépolarisation ventriculaire.
• Le complexe QRS représente la dépolarisation ventriculaire.
• L'onde T représente la repolarisation ventriculaire.
La repolarisation auriculaire est masquée par la dépolarisation ventriculaire.

SYSTÈME CARDIOVASCULAIRE – Physiologie cardio-vasculaire
5/16
e) La dépolarisation cardiomyocytaire
On prend en modèle le myocyte ventriculaire standard, donc la cellule musculaire et la cellule du tissu
nodal.
On représente le potentiel membranaire en fonction du temps.
Phase 0 : Dépolarisation rapide
Phase 1 : Repolarisation rapide partielle
Phase 2 : Plateau de dépolarisation
Phase 3 : Repolarisation lente
Phase 4 : Potentiel de repos
Cette dépolarisation est due à des phénomènes ioniques dans la cellule.
La phase 0, ou dépolarisation rapide, est due à un flux sodique rapide entrant très important du fait de
l'augmentation de la conductance des canaux sodiques.
La phase 1, ou repolarisation rapide partielle, est due à modification de la perméabilité d'un canal potassique
rapide. C'est la sortie de potassium qui provoque ce début de repolarisation.
La phase 2, ou plateau de repolarisation, est due à l'entrée de calcium provoquée par le canal calcique lent de
type L. Celui-ci provoque une entrée de Ca2+, qui contrebalance l'action de la sortie de K+ et est à l'origine du
plateau de repolarisation.
La phase 3, ou repolarisation lente, est due à la baisse de conductance du canal calcique et donc de la
diminution du flux entrant de calcium, et aux canaux potassiques lents qui provoquent la sortie de
potassium.
La phase 4 caractérise le retour au potentiel de repos.
La dépolarisation est donc un phénomène électrique sous la dépendance de phénomènes ioniques dont les
différents acteurs sont le calcium, le sodium et le potassium.
Pour rappel la concentration en sodium est 30 fois plus importante en extracellulaire ce qui favorise son
entrée dans la cellule. C'est l'inverse pour le potassium qui est 30 fois plus concentré en intracellulaire, ce
qui favorise sa sortie.
 6
7
8
9
10
11
12
13
14
15
16
6
7
8
9
10
11
12
13
14
15
16
1
/
16
100%