23/02/15 DROUILLY Marie L3 CR: MACIOW Benjamin

SYSTÈME CARDIOVASCULAIRE – Physiologie cardio-vasculaire (1)
23/02/15
DROUILLY Marie L3
CR: MACIOW Benjamin
Système cardiovasculaire
Pr Stéphane Delliaux
18 pages
Physiologie cardio-vasculaire (1)
A. Introduction-Généralités
Andrea Cesalpino (1519-1603) a défini le terme de circulation, il en attribue le rôle au cœur et introduit la
notion de mouvement perpétuel.
La fonction du cœur est de mobiliser le sang et de permettre la circulation. Ceci afin de transporter l'O2 et les
nutriments vers les tissus, puis de collecter le CO2 et les produits du métabolisme afin de les acheminer vers les
organes épurateurs (rein, foie, poumons).
La machinerie cardiaque est un moteur bien huilé, on compte environ 60 battements par minute soit
2,4.109 battements tout au long de la vie.
C'est un organe vital. Lorsqu'il est défaillant, cela aboutit à des pathologies cardiaques à l'origine d'une
mortalité non négligeable. C'est un organe autonome avec une activité propre mais qui est orchestré par le
système nerveux végétatif.
C'est un organe précieux, nécessaire, hautement spécialisé composé de plusieurs tissus. Il est composé :
-essentiellement de muscle strié viscoélastique et contractile. C'est un organe excitable, il se comporte comme
un syncytium: une cellule transmet son état électrique, ionique, métabolique aux cellules adjacentes qui se
mettent du coup dans une même condition cellulaire permettant une uniformité de fonction.
Le cœur a une activité contractile autonome=myogénique (naît dans le muscle lui même) et rythmique.
-mais aussi de tissu fibreux qui est plutôt structurant (anneaux valvulaire) mais surtout électrogénique
1/18
Plan
A. Introduction-Généralités
B. La contraction cardiaque
I. L'électrogénèse cardiaque
II. Le couplage excitation-contraction
III. Le raccourcissement cardiomyocytaire
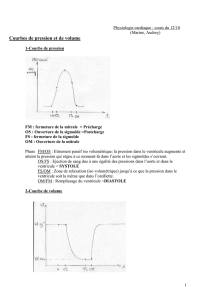
C. Le cycle cardiaque
I. Généralités
II. Le cycle diastole-systole
III. Le diagramme de Wiggers
D. Énergétique cardiaque
I. Consommation cardiaque en oxygène
II. La contraction non chargée
III. La contraction chargée

SYSTÈME CARDIOVASCULAIRE – Physiologie cardio-vasculaire (1)
B. La contraction cardiaque
I. L'électrogenèse cardiaque
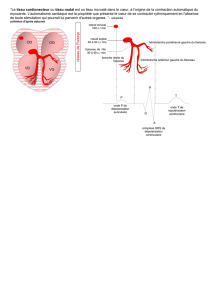
a) Le tissu nodal
Ce sont des cellules myocardiques qui ont gardé certaines propriétés des cellules embryonnaires, elles ont donc
des propriétés particulières:
-automaticité: elles ont une activité propre
-rythmicité: cette automaticité va se répéter dans le temps de manière cyclique
-excitabilité: capacité à générer une activité électrique et à se dépolariser
-auto-excitabilité: la cellule produit elle-même un phénomène de dépolarisation indépendamment du milieu
extérieur
-conductibilité
-contractilité: due aux cellules myocardiques
Le tissu nodal permet la genèse d'une activité électrique.
Le tissu nodal comprend le nœud sinusal, le nœud atrio-ventriculaire et des faisceaux (tronc du faisceau de
His et ses ramifications, les faisceaux inter atrio-ventriculaire, réseau de Purkinje).
b) L'auto-excitabilité du tissu nodal
L'auto-excitabilité est une propriété particulière des cellules du tissu nodal, correpond à leur capacité de générer
un phénomène électrique d'elle-même, sans signaux extérieurs.
La cellule ventriculaire n'appartenant pas au tissu nodal, elle a un potentiel de repos (de -90mV) stable et
constant au cours du temps jusqu'à la survenue d'un phénomène électrique avec une dépolarisation stéréotypée
puis un retour au repos.
2/18

SYSTÈME CARDIOVASCULAIRE – Physiologie cardio-vasculaire (1)
Pour la cellule sinusale, on s'aperçoit que le potentiel de repos est moins électronégatif (-60mV) et surtout
qu'il n'est pas constant puisque il a tendance à décroître au cours du temps jusqu'à ce qu'il dépasse le seuil
d’excitabilité puis va être à l'origine d'un potentiel d'action, puis retour à un potentiel de repos instable,...
La tendance spontanée à la dépolarisation entraîne une contraction rythmique.
c) La dépolarisation diastolique lente
Au cours de la diastole, il y a une dépolarisation lente des cellules du tissu nodal due à des phénomènes
ioniques.
Elle est due à la conjoncture d'au moins 3 phénomènes :
- le courant de fond
- le pace-maker
- le courant calcique transitoire
Le courant de fond est un échange qui se fait grâce à la pompe électrogénique Na+/K+ ATPase et à un
échangeur Na+/Ca2+ qui fait rentrer du sodium et sortir du calcium.
Dans la dépolarisation diastolique lente, au niveau du courant de fond il y a une diminution de la pompe
électrogénique qui entraîne une accumulation sodique intracellulaire et une hyper-activation de l'échangeur
majorant encore le flux sodique entrant.
Le courant pace-maker est dû à des canaux ionique HCN qui sont des canaux sodiques dépendant du degré
d'hyper-polarisation de la cellule. Plus la cellule est électronégative, plus ce canal ionique va être actif et va
générer un flux entrant de sodium ce qui va favoriser la dépolarisation.
Le flux calcique transitoire est assuré par un canal calcique de type T voltage-dépendant. L'augmentation
spontanée du potentiel de membrane qui va tendre vers 0 va être activée par le phénomène de dépolarisation et
génère un flux calcique entrant.
La conjoncture de ces trois phénomènes ioniques est à l'origine de la dépolarisation spontanée diastolique
lente.
3/18

SYSTÈME CARDIOVASCULAIRE – Physiologie cardio-vasculaire (1)
L'ensemble de ces canaux ioniques a une multitude d'expressions phénotypiques différentes. Ainsi dans le nœud
sinusal, le nœud atrio-ventriculaire, le faisceau de His,... on a des phénomènes électriques de morphologie
légèrement différente.
Le tissu sinusal a la dépolarisation la plus rapide, c'est celui qui va se dépolariser le plus rapidement et à une
fréquence plus élevée. Il va donc imposer un certain rythme, le rythme sinusal, de l'ordre de 60 bpm.
S'il ne fonctionne plus, la deuxième structure qui a la fréquence la plus rapide prendra le pas, c'est le rythme
jonctionnel (30-40bpm) au niveau du nœud atrio-ventriculaire. Si celui-ci n'est plus fonctionnel, on a encore
d'autres structures comme le rythme idio-ventriculaire (10-20 bpm).
L'ensemble de ces phénomènes électrique quand on les somme dans le temps et l'espace, donne l'ECG avec
l'onde P, le complexe QRS et l'onde T.
d) La conduction de l'excitation cardiaque
Quand la dépolarisation naît, il y a un début d'excitation puis une propagation de ce phénomène électrique pour
avoir une contraction. C'est la conduction qui part du nœud sinusal, se répand dans les oreillettes avant d'aller
dans le secteur ventriculaire en passant par le nœud atrio-ventriculaire et le faisceau de His, puis une
dépolarisation ventriculaire qui naît à l'apex et remonte dans l'ensemble des cellules myocardiques.
La vitesse de conduction est très augmentée dans le faisceau de His et dans l'apex, ils sont donc excités avant le
reste du ventricule.
4/18

SYSTÈME CARDIOVASCULAIRE – Physiologie cardio-vasculaire (1)
Le début de la dépolarisation qui naît dans le nœud sinusal correspond au début de l'onde P, la propagation de
l’excitation aux 2 oreillettes correspond à l'onde P dans sa totalité, l'excitation ventriculaire et de l'apex c'est Q
et la généralisation du phénomène électrique à l'ensemble du myocarde correspond au complexe QRS.
La repolarisation correspond à l'onde T. Elle suit le même principe, elle naît au niveau de l'apex et se termine au
niveau du ventricule.
e) La dépolarisation cardiomyocytaire
La courbe la plus haute représente un potentiel d'action cardiomyocytaire classique avec 5 phases :
Phase 0: dépolarisation rapide, complète et maximale à partir d'un potentiel de repos constant, -90mV
Phase 1: repolarisation rapide partielle
Phase 2: plateau de dépolarisation
Phase 3: repolarisation lente en plusieurs dizaines voire centaines de millisecondes
Phase 4 : potentiel de repos
Ce phénomène électrique est dû à des phénomènes ioniques.
Il y a 3 phénomènes ioniques majeurs dont la conjonction est à l'origine de la repolarisation rapide partielle. :
• Modification de l'homéostasie sodique: augmentation très rapide, importante et brève de la
conductance sodique, à l'origine d'un flux sodique rapide entrant. C'est ce phénomène ionique qui est à
l'origine de la dépolarisation rapide.
• U n courant calcique entrant survient quelques millisecondes plus tard. Il est assurée par un canal
calcique lent de type L. On a un flux calcique entrant lent qui dure tout le potentiel d'action, il y a un premier
pic, un plateau puis il décroit. Il est à l'origine du plateau de dépolarisation.
• Phénomène potassique rapide sous-tendu par la conductance de canaux potassiques qui décroit de
façon très rapide pendant les premières dizaines de millisecondes, puis augmentation progressive afin
d'atteindre ses valeurs de repos à la fin du phénomène électrique. C'est cette modification progressive de la
conductance potassique qui est à l'origine de la repolarisation lente et du retour au potentiel de repos.
5/18
 6
7
8
9
10
11
12
13
14
15
16
17
18
6
7
8
9
10
11
12
13
14
15
16
17
18
1
/
18
100%