TH1_ch3

TH1_ch3
La contraction musculaire, un couplage
chimio mécanique impliquant des
protéines motrices

Coupe longitudinale; taille cellulaire:30 cm x 100 µm Coupe transversale
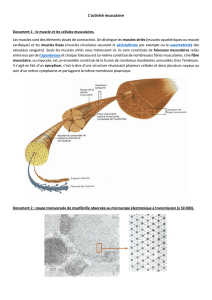
cellules très allongées et
cylindriques avec de
nombreux noyaux Cellule
polynucléé

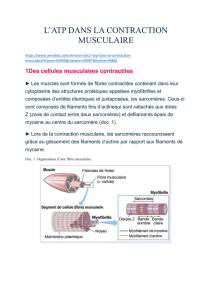
Un muscle = X fibres (cellules)
musculaires = X myofibrilles
[striation longitudinale] = X
myofilaments protéiques d’actine
et de myosine

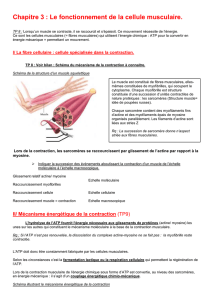
Chaque myofibrille est entourée par du
reticulum sarcoplasmique (reticulum
endoplasmique musculaire) selon une
organisation rigoureuse. A intervalles plus
ou moins réguliers, le reticulum
sarcoplasmique émet des protubérances
appelées citernes terminales.
A ces niveaux se trouvent également des
invaginations de la membrane
cytoplasmique appelées tubules
transverses.
On trouve donc un tubule transverse en
étroite association avec deux citernes
terminales, formant ce que l'on appelle une
triade (Figure 2). Cette organisation joue un
rôle important dans le couplage excitation-
contraction

 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
1
/
20
100%