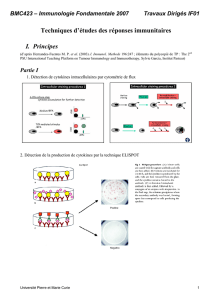
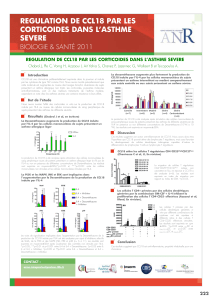
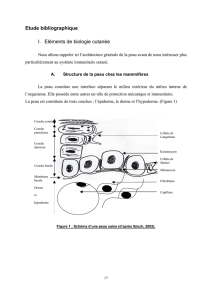
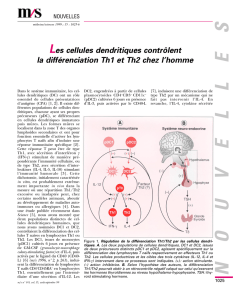
Interleukine 4 - iPubli

SYNTHÈSE
Jacques Banchereau
27,
69570
Interleukine

1 Sources ceulaires
de lnterleukine
1
2
3

1989.)

�
°
·

l
T
 6
7
8
6
7
8
1
/
8
100%