Immunité et grossesse - iPubli

SYNTHÈSE
Directeur de recherche au Cnrs
Immunité et grossesse
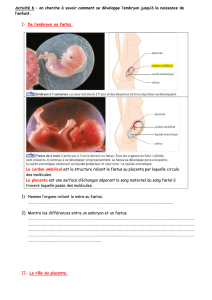
igures 1 et 2, page
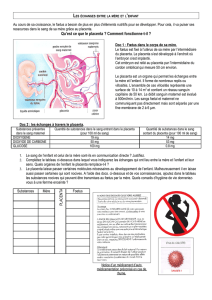
gure page


-
1

1

A
8
 6
7
8
9
6
7
8
9
1
/
9
100%