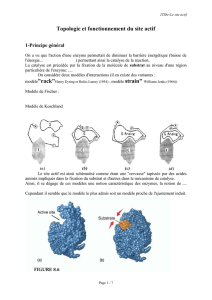
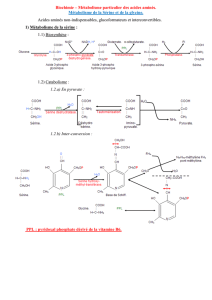
10 scimd met aa nov 2012

1
Métabolisme de composés azotés
INTRODUCTION
Les composés azotés sont des composants essentiels de la plupart des organismes vivants. Parmi ces composés les
acides aminés occupent une position centrale, notamment du fait de leur rôle amphibolique :
- incorporation dans la genèse de novo des nucléotides puriques ou pyrimidiques
- synthèse des porphyrines
- synthèse des hormones peptidiques…
I. REACTIONS GENERALES DES AMINOACIDES
I. 1. Réactions enzymatiques impliquant le phosphate de pyridoxal (PAL)
Le PAL est impliqué dans un grand nombre de réactions intéressant le métabolisme des AA (décarboxylation,
transamination, racémisation entre autres. Il ne peut être synthétisé dans notre organisme d’où l’activité
vitaminique B6 manifestée tant par le pyridoxal que par la pyridoxine (l’alcool primaire correspondant) et par la
pyridoxamine (l’amine primaire correspondante, qui se forme d’ailleurs intermédiairement lors du processus de
transamination).
A.

2
Décarboxylation
La décarboxylation des AA conduit à une amine. Certaines de ces amines ont un rôle physiologique ou
pharmacologique important, d’où le nom d’amines biogènes qu’on leur donne parfois.
Les décarboxylations sont catalysées par des décarboxylases, enzymes à phosphate de pyridoxal, présentes chez
les micro-organismes et dans les tissus animaux. Les bactéries intestinales ont des enzymes capables de
décarboxyler la lysine en cadavérine et l’ornithine en putrescine. Ces amines sont toujours présentes en petites
quantités dans l’intestin, mais qui peuvent provoquer des intoxications si leur concentration augmente à la suite
de fermentations intestinales anormales.
Principales amines biogènes.
Cystéine
mercaptoéthylamine
dans le coenzyme A
Histidine
histamine
action hypotensive
3,4- dihydroxy-
phénylalanine (DOPA)
.
dopamine
precurseur de l’adrenaline
Sérine
éthanolamine
dans les phosphatides
5-hydroxy-tryptophane
sérotonine
hormone tissulaire vasoconstrictrice
B. Transamination
Ce terme désigne le transfert réversible du groupement amine d’un aminoacide à un -cétoacide; il n’y a pas
libération de NH3, contrairement à ce qui se passe lors de la désamination.

3
Les transaminases sont universellement répandues chez les bactéries, les végétaux et les animaux. Il existe un
grand nombre de transaminases, spécifiques des divers aminoacides, mais il y en a deux qui sont particulièrement
abondantes dans les tissus animaux, et qui ont été bien étudiées; elles catalysent les deux réactions suivantes
acide L-glutamique + acide oxalo-acétique ↔ acide a-cétoglutarique + acide L-aspartique
Transaminase Glutamate Oxaloacétate (TGO) ou Aspartate Aminotransférase (ASAT)
acide L-glutamique + acide pyruvique ↔ acide a-cétoglutarique + L-alanine
Transaminase Glutamate Pyruvate (TGP) ou Alanine aminotransférase
Dans les deux cas, le groupement aminé de l’acide glutamique est transféré à un -cétoacide pour former
l’aminoacide correspondant.
II. 2. La désamination
A. La désamination oxydative
Ce processus essentiellement irréversible a lieu en deux étapes. Dans un premier temps, l’enzyme catalyse une
déshydrogénation de l’aminoacide en iminoacide, cette enzyme est une flavoprotéine et les 2 électrons enlevés au
substrat viennent réduire le FAD en FADH2 (ou le FMN en FMNH2). L’iminoacide est spontanément hydrolysé en
-cétoacide + ammoniac.

4
L’acide glutamique est désaminé par une enzyme particulière, la L-glutamate-déshydrogénase, enzyme à NAD ou
à NADP selon les organismes. Cette réaction, contrairement aux autres désaminations oxydatives, est réversible, et
c’est dans le sens de l’amination réductrice qu’elle est particulièrement importante. C’est de ce fait l’un des
principaux processus de fixation de l’ammoniac dans les composés organiques, et le groupement aminé ainsi
formé pourra aisément passer de l’acide glutamique aux autres aminoacides par transamination. En outre, ce
passage de l’acide glutamique à l’acide a-cétoglutarique (qui est l’un des composés du cycle de Krebs) et vice versa
constitue l’un des points de contact entre les métabolismes glucidique et protéique.
Chez les animaux supérieurs (Mammifères), la glutamate déshydrogénase a un rôle surtout catabolique. L’enzyme
est mitochondriale et fonctionne dans le sens de la désamination oxydative avec le NAD, en association avec les
transamjnases pour cataboliser les excédents d’aminoacides. Dans les mitochondrjes hépatiques, l’ammoniaque
produit par l’enzyme sera à l’origine de la synthèse de l’urée (voir cycle de l’urée).
B. Désamination désaturante
La désamination ne concerne qu’un petit nombre d’aminoacides. Dans ce processus, l’enzyme catalyse
l’enlèvement d’une molécule d’ammoniac avec formation d’une double liaison entre les carbones et . Par
exemple la désamination de l’acide L-aspartique en acide fumarique, réaction réversible qui permet la fixation de
NH3 dans un composé du cycle de Krebs. Cette réaction est catalysée par l’aspartate-ammonium lyase, présente
chez les micro-organismes et les végétaux. C’est selon ce mode sont également désaminées l’histidine chez les
animaux et la phénylalanine chez les végétaux.

5
II. ORIGINE DES AMINOACIDES DANS LES ORGANISMES VIVANTS
Concernant l’origine des aminoacides, les organismes vivants se divisent en deux groupes :
- ceux qui sont capables de synthétiser tous les vingt aminoacides naturels, et chez lesquels les aminoacides sont
donc d’origine endogène
- et ceux qui sont incapables de synthétiser certains aminoacides; ces aminoacides qui leur sont indispensables,
doivent leur être fournis dans le milieu (ou dans l’alimentation) et sont donc d’origine exogène.
1. Synthèse des aminoacides
La synthèse des aminoacides peut être subdivisée en plusieurs étapes : la formation d’ammoniac, l’incorporation
d’ammoniac dans un composé organique (en général sous forme d’acide glutamique), la synthèse du squelette
carboné des aminoacides (c’est-à-dire la formation des -cétoacides correspondants) et enfin le transfert du
groupement aminé de l’acide glutamique à ces divers -cétoacides par transamination.
A. Formation d’ammoniac
La plupart des micro-organismes et des végétaux utilisent l’ammoniac qui se forme lors de la dégradation des
substances organiques azotées. On peut également observer la formation d’ammoniac à partir de substances
minérales : l’azote atmosphérique et les ions nitrite et nitrate par réduction.
B. Incorporation de l’ammoniac dans un composé organique
Les quatre processus les plus importants permettant la « fixation » de NH3 sous forme organique sont :
- l’amination réductrice de l’acide -cétoglutarique en acide glutamique par la glutamate DH;
- l’amination de l’acide fumarique en acide aspartique ;
- la formation de carbamyl-phosphate ;
- la formation d’une amide comme la glutamine.
Les divers modes de formation de l’ammoniac et son incorporation dans les composés organiques peuvent être
considérés dans le cadre général du cycle de l’azote.
C. Synthèse du squelette carboné des aminoacides
Les -cétoacides correspondant à un certain nombre d’aminoacides sont des produits courants du métabolisme
intermédiaire, notamment de la glycolyse et du cycle de Krebs; c’est le cas particulier de l’acide pyruvique
(-cétoacide correspondant à l’alanine), de l’acide -cétoglutarique (correspondant à l’acide glutamique), de
l’acide oxalo-acétique (correspondant à l’acide aspartique). Certains autres -cétoacides dérivent plus ou moins
directement de composés du métabolisme intermédiaire, comme par exemple l’acide 3-phospho-hydroxy-
pyruvique (correspondant à la phospho-sérine) qui provient de l’acide 3-phospho-glycérique par
déshydrogénation. Mais alors que les micro-organismes et les végétaux sont capables de former une dizaine
d’-cétoacides, les animaux supérieurs et l’Homme en particulier sont incapables de réaliser la synthèse de la
majorité d’entre eux et il faut leur fournir dans l’alimentation les aminoacides correspondants, ce sont les
aminoacides « indispensables ». En ce qui concerne les organismes capables de synthétiser cette dizaine d’-
cétoacides, les réactions nécessaires sont parfois nombreuses et complexes, notamment pour les aminoacides
aromatiques..
D. Transfert du groupement aminé sur les -cétoacides
De façon générale, les aminoacides peuvent être formés à partir des -cétoacides correspondants, par
transamination. Comme l’acide glutamique est le principal composé résultant de l’incorporation de NH3 sous forme
 6
7
8
9
10
11
12
13
6
7
8
9
10
11
12
13
1
/
13
100%