Bioénergétique de la contraction musculaire

582663756 1/16
B
BI
IO
OE
EN
NE
ER
RG
GE
ET
TI
IQ
QU
UE
E
D
DE
E
L
LA
A
C
CO
ON
NT
TR
RA
AC
CT
TI
IO
ON
N
M
MU
US
SC
CU
UL
LA
AI
IR
RE
E
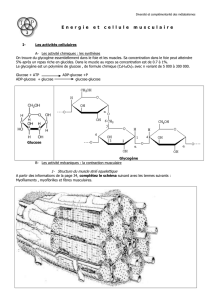
La contraction musculaire revient à l’hydrolyse de l’ATP (ATP ADP + Pi + énergie).
L’énergie chimique produite par l’hydrolyse de l’ATP va être transformée en énergie
mécanique.
I. ORIGINE DE LA CONTRACTION
contraction musculaire = glissement de la myosine sur l’actine
diminution de la longueur du sarcomère (= zone limitée par 2 bandes H)
raccourcissement du muscle
A. AU REPOS
Schéma : Bases structurales et fonctionnelles
La tête de myosine est maintenue à distance de l’actine. Il n’y a pas contact entre l’ATP et
son site sur l’actine car les molécules de Tropomyosine s’interposent.
La tête de myosine présente une activité ATPase masquée. Cette activité ATPasique sera
dévoilée lors de la liaison actine-myosine.

582663756 2/16
L’ATP est l’élément chimique qui permet la contraction musculaire, mais
l’hydrolyse de l’ATP n’est pas possible au repos car la Tropomyosine empêche le
contact entre la tête de myosine et l’actine.
B. LORS DE LA CONTRACTION
Schémas : Résumé des mécanismes de la contraction musculaire

582663756 3/16
Le potentiel d’action neuromusculaire provoque l’arrivée du Ca2+ aux myofibrilles.
Le Ca2+ va se fixer sur la Troponine C.
conséquences : déplacement de la troponine
liaison actine-myosine
action ATPase dévoilée levée
hydrolyse de l’ATP
contraction = Inclinaison des têtes de myosine
diminution de l’angle qu’elle fait au repos avec la myosine (de 90° à 45°)
L’hydrolyse de l’ATP est la transformation de l’énergie chimique en énergie mécanique ;
c’est l’élément qui provoque la contraction.
Schéma : La molécule d’ATP (Adénosine Triphosphate)
Schéma : L’hydrolyse de l’ATP
chaleur
ATP + H2O ADP + Pi + énergie libre
travail musculaire

582663756 4/16
Cependant, la concentration musculaire en ATP est faible (~ 5 mmoles/kg), ce qui ne
permet d’effectuer des contractions que de quelques secondes.
Pour que le muscle se contracte plus longtemps, il faut un apport suffisant en ATP ; il faut
donc en resynthétiser. C’est là qu’interviennent les substrats énergétiques et les différentes
voies métaboliques de dégradation de ces substrats.
L’intervention des différentes voies métaboliques est conditionnée par la durée et
l’intensité de l’effort. Cette intervention permet la resynthèse de l’ATP (qui est nécessaire
à la contraction musculaire mais qui est présent en faible quantité dans le muscle).
Selon la voie métabolique, certains substrats vont être utilisés préférentiellement (ex : les
lipides pour les exercices de longue durée et de faible intensité ( aérobie)).
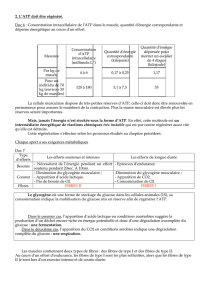
II. SUBSTRATS ENERGETIQUES
Tableau : Substrats disponibles chez un homme de 80kg ayant 25kg de muscles.
Substrats Quantité Energie disponible
(en kg) (en kJ)
Triglycérides 12 446 500
Protéines (acides aminés libres) 6 78 250
Glycogène
hépatique 0,060 638
musculaire 0,400 4252
Substrats circulants 0,023 420
(glucose, acides gras)
Phosphorylcréatine 0,087 17
ATP 0,076 3
rq : Il faut éviter de faire des efforts de trop longue durée sous peine de puiser dans les
réserves de protéines et de perdre ainsi de la masse musculaire.
L’utilisation des substrats est fonction de l’intensité et de la durée de l’exercice.

582663756 5/16
III. PROCESSUS METABOLIQUES
La resynthèse de l’ATP nécessite l’intervention des processus métaboliques :
aérobie (présence d’oxygène) exercice de longue durée et de faible ou moyenne
intensité
anaérobie (absence ou manque d’oxygène) exercice de courte durée et de haute
intensité
Il existe un temps d’adaptation de la fonction respiratoire à l’exercice.
Pour un exercice d’une durée de 2 à 3 min, l’oxygène est très peu utilisé pour procurer de
l’énergie. Ce sont donc les processus anaérobies qui sont principalement utilisés.
A. PROCESSUS METABOLIQUES ANAEROBIES
Anaérobie alactique
- absence d’O2
- pas de production d’acide lactique
- intervient dès le début de l’exercice
- efforts de 3 à 4’’ à intensité quasi-maximale
Anaérobie lactique
- manque d’O2
- production d’acide lactique
- effort de haute intensité prolongé
1. ANAEROBIE ALACTIQUE
La chute de concentration en ATP est évitée par 2 mécanismes :
myokinase
1er mécanisme : 2ADP AMP + ATP ([ATP] = 10-3 mmol/kg)
Cette réaction est rendue possible grâce à une enzyme : la Myokinase.
L’ADP étant présent en quantité négligeable, ce mécanisme ne va pouvoir assurer que
quelques contractions musculaires.
 6
7
8
9
10
11
12
13
14
15
16
6
7
8
9
10
11
12
13
14
15
16
1
/
16
100%