Chapitre 2 :

Chapitre 2 :
- Du carbone minéral aux composants du vivant : La photo-autotrophie pour le Carbone -
Les végétaux sont photo-autotrophes pour le carbone : ils élaborent leur matière organique grâce à l’énergie
lumineuse. La transformation du carbone minéral en carbone organique est permise grâce au processus de la
photosynthèse qui se déroule dans les chloroplastes. Ce processus nécessite donc de la lumière, de la chlorophylle, du
CO2 et de l’eau. Elle produit des glucides et un déchet, le dioxygène.
Comment le carbone minéral du CO2 est-il fixé et réduit dans les molécules organiques ?
Comment l’énergie nécessaire à cette fixation est-elle fournie par la lumière ?
1°)- La photosynthèse : Une réaction en deux étapes :
Livre p.47 – Questions 1,2 et 3
Deux étapes complémentaires et étroitement liées en deux régions du chloroplaste :
- La phase photochimique dans la membrane des thylakoïdes permet la conversion de l’énergie lumineuse en
énergie chimique.
- La phase non photochimique (ou thermochimique) dans le stroma et où se réalise l’assimilation du CO2 grâce à
l’énergie chimique.
2°)- La capture de l'énergie lumineuse :
2-1- La lumière= source d'énergie :
Comment l’énergie lumineuse est-elle
réceptionnée par les chloroplastes ?
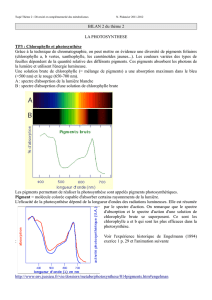
Etude des pigments chlorophylliens par
chromatographie (Doc1 p.48) ; spectroscopie
manuelle et en exao.
Q1- A partir de l’exploitation de la
chromatographie, du document 3 page 49 et du
document ci-dessous, préciser comment le
chloroplaste réceptionne la lumière.
Q2- Rajouter sur le graphique ci-dessous,
l’allure probable de la courbe correspondant à
l’évolution de l’activité photosynthétique en
fonction de la longueur d’onde. Justifier cette
évolution.
La lumière blanche mélange de radiations lumineuses de λ (longueur d’onde) visibles comprises entre 400 et 700 nm
permet la photosynthèse chez les végétaux chlorophylliens car chaque λ véhicule de l’énergie dans ses photons (E = h C/ λ)
L’intensité de la photosynthèse d’une plante chlorophyllienne varie selon la nature de l’éclairement : elle est maximale
dans les radiations bleue-violette, faible dans le vert-jaune et nulle dans le rouge foncé =spectre d’action
La chlorophylle brute extraite des végétaux contenant des pigments tels que la chlorophylle a, b,
les xanthophylles et
le carotène,
absorbent plus ou moins les différentes longueurs d’onde sauf le vert (d’où la couleur verte des
végétaux)
Le spectre d’action de la chlorophylle brute concorde avec le spectre d’absorption du végétal : les radiations
les mieux absorbées sont les plus efficaces pour la photosynthèse.
2-2- Les photosystèmes du Chloroplaste :
Schéma bilan Chloroplaste (Q3 p.47)
Les pigments situés dans la membrane des thylakoïdes des chloroplastes sont des molécules capables d’absorber
certaines radiations lumineuses et piègent l’énergie lumineuse (capture de photons) qui permet la photosynthèse.
Ils sont donc appelés pigments chlorophylliens. Ils sont associés à des protéines de la membrane des thylakoïdes et
constituent des complexes ou photosystèmes (doc4 p.51)

3°)- Da la lumière à l'ATP : La phase photochimique : Voir TP5 + Doc.C p.51
Dans les thylakoïdes, grâce à la collecte des photons par les photosystèmes, un ensemble d’oxydoréductions permet
l’oxydation de l’eau, la production d’O2 et de composés intermédiaires (coenzymes réduits) RH2 et d’ATP (adénosine
triphosphate qui se construit à partir d’ADP et de phosphate inorganique).
Des transporteurs d’électrons sont donc réduits et une enzyme insérée dans la membrane, l’ATP syntéthase , permet
la production d’ATP à partir de phosphorylation d’ADP (ADP+Pi → ATP).
La chlorophylle excitée dans la membrane des thylakoïdes perd ses électrons captés par une chaîne de transporteur d’électrons dans la
membrane des thylakoïde. Ils sont transferés jusqu’à un accepteur final R+ situé dans le stroma :
R+ + 2H+ + 2 e-
RH2 (coenzyme réduit à fort pouvoir réducteur susceptible d’être transmis sur d’autres moléculesultérieurement).
La régénération de la chlorophylle oxydée, c'est-à-dire sa réduction, est assurée par une oxydo-réduction avec l’eau : H2O oxydée
fournit les e- réduisant la chlorophylle. La photolyse de l’eau libère aussi O2,et des protons H+. Ceux-ci se concentrent à l’intérieur des
thylakoïdes et crée une différence de pH entre lumen (intérieur des thylakoïdes et stroma). Ce gradient de pH qui s’installe à travers la
membrane du thylakoïde représente une énergie potentielle qui permet la synthèse d’ATP (Adénosine Tri Phosphate) au niveau de
sphères pédonculées de la membrane : les ATP synthétases.
L’ATP est une petite molécule hautement énergétique disponible au niveau du stroma dont l’hydrolyse libère une quantité d’énergie
utilisée par de nombreuses réactions enzymatiques.
4°)- La fixation et la réduction du dioxyde de carbone : La phase non photochimique
Livre Unité 4 p.52, 53 questions 1,2 et3.
*Objectifs : On cherche à comprendre
L’enchaînement des réactions permettant la fixation et la réduction du CO2 – Doc.1,2 et 3
La régénération de l’accepteur du carbone et donc le caractère cyclique de l’assimilation-
Doc.3
Le rôle des intermédiaires chimiques issus de la phase photochimique (RH2 et ATP).
*Document 1 p.52 : Description de l’expérience de Calvin et explication du principe.
- Des Chlorelles en présence de lumière + CO2 réalise la photosynthèse ;
- Le 14CO2 permet de marquer les produits intervenant dans l’assimilation du CO2 ;
- Le tubulure transparent permet au Chlorelle de poursuivre la photosynthèse sortie
du réservoir en présence de 14CO2 fourni à différents niveaux avant de les fixer
dans l’alcool bouillant. Ainsi on peut suivre cette assimilation au cours du temps.
*Document 2 : Chromatographie bidirectionnelle ( ? à expliquer ?) puis autoradiographie
*Question 1 p.53 : Nature de la première molécule formée lors de la fixation du CO2 – Doc.2a :
Après 5sec de photosynthèse en présence de 14 CO2 on peut retrouver la radioactivité la plus
importante dans une molécule de PGA, une molécule en C3 qui n’est pas un glucide (C3P)
Donc le CO2 est fixée en premier dans du PGA (Phosphoglycérate ou acide phosphoglycérique).
*Question 2 p.53 : Nature de l’accepteur de CO2 –
Doc.3a :
Lorsque l’on suit les taux de PGA et DE C5P2, on observe qu’en présence de lumière leur
taux son constant puis en son absence, les taux s’inversent : augmentation des quantités de PGA
et diminution de celles de C5P2 jusqu’à se stabiliser.

Il semblerait donc que le PGA se forme au dépend du C5P2 lors de la phase non photochimique !
Doc.3b :
Par contre en l’absence de CO2 (ou plutôt en concentration très faible), les évolutions
sont inversées : Augmentation du C5P2 et parallèlement diminution du PGA.
Ainsi la faible concentration de CO2 ne permet plus la synthèse de PGA, libérant le C5P2 de sa
participation à cette réaction.
Il faut donc admettre que l’accepteur de CO2 est C5P2, un glucide en C5 (doc2) formant ensuite
du PGA suivant la réaction : CO2 + C5P2 C6P2 2 molécules de PGA (en C3)
sans relation directe avec la lumière mais nécessitant les intermédiaires chimiques RH2 et ATP.
Q2 (suite) : Le devenir des produits formés –
Doc.2b :
30 sec. de photosynthèse en présence de 14 CO2 permet donc de retrouver le CO2 fixé
non seulement dans le PGA mais aussi le C5P2, le C3P (glucide phosphorylés à 3 carbones) et
d’autres sucres P, acides aminés…
Doc.4 :
Si l’on suit l’évolution en parallèle du PGA et des sucres P, on remarque que l’un dépend de
l’autre : Le PGA permet de former les sucres P.
Donc on peut admettre aussi que le PGA formé à la suite de la fixation du CO2 formera à son tour
du C3P, des sucres P, des acides aminés (grâce à l’apport de l’azote, phosphore…provenant de la
sève brute).
Q2 (suite) : Cette assimilation du CO2 est cyclique –
Doc.3b :
Apparition du C5P2 dans le chromatogramme. Ce précurseur du PGA finit par radioactif
au bout de 30 sec.. Ainsi le PGA formé doit participer à la régénération du C5P2 pour que la
fixation du CO2 se poursuive.
Doc.4a :
En phase lumineuse, les taux de PGA et C5P2 sont constants. Donc les concentrations
des deux composés sont équilibres. En absence de lumière, on crée un déséquilibre par l’arrêt de
la phase photochimique : le C5P2 n’est plus renouvelé et la fixation du CO2 finira pas ne plus se
faire (stagnation du PGA)
Ainsi le C5P2 ne peut se régénérer que grâce à la phase photochimique grâce à l’apport des
intermédiaires chimiques : RH2 et ATP et avec la participation d’une partie du PGA (schéma)
Dans le stroma, il y a incorporation et réduction du CO2 pour la synthèse des glucides (CH2O). Cela nécessite un
accepteur de CO2, de l’ATP, et des composés réduits RH2.
Il y a un cycle de réduction photosynthétique du carbone.
Le CO2 est fixé par un accepteur organique à 5C, le ribulose 1-5 bi phosphate ou C5P2 ; la réaction est catalysée
par une carboxylase présente dans le stroma au cours de la photosynthèse.
Cette réaction de carboxylation, non photochimique (qui peut s’effectuer à l’obscurité), dépendante de la
température, conduit à la formation de 2 molécules à 3 carbones, le phosphoglycérate ou acide phosphoglycérique
APG
Il y a couplage entre la fixation de CO2 et la phase photochimique : les molécules en C3 formées par la carboxylase
sont transformées en molécules glucidiques, les trioses phosphates (2C3P). Cette réaction nécessite de l’ATP et des
coenzymes réduits - ou RH2 - produits par les réactions photochimiques dans les thylakoïdes des chloroplastes
éclairés. Ainsi, bien que cette phase non photochimique ne nécessite pas directement de la lumière, elle reste
entièrement dépendante des produits de la phase photochimique.
De nombreuses réactions biochimiques catalysées par des enzymes permettent de réaliser de nombreuses molécules
organiques à partir des trioses phosphates : glucides, acides gras, acides aminés (avec apport de N et de S).
Une partie des trioses phosphates permet de reconstituer le stock d’accepteurs de CO2 (C5P2) indispensables à
l’incorporation de CO2 . Le processus non photochimique est donc cyclique (cycle de Calvin).

Les molécules synthétisées peuvent être stockées dans le chloroplaste sous forme d’amidon, exportées hors du
chloroplaste, distribuées dans la plante par les tubes criblés du liber sous forme de saccharose pour être utilisées
par les cellules sous forme de glucose ou fructose comme source d’énergie et de matière première (respiration et
synthèse).
Bilan : La photosynthèse apparaît donc comme la voie d’entrée du carbone minéral et de l’énergie solaire dans
la biosphère et est donc à l'origine de l'ensemble de la biomasse.
Voir le document associé au TP5 :
Reprendre le croquis des réactions photochimiques afin de l'inclure dans un plus
vaste schéma à l'échelle d'un chloroplaste (avec sa légende) afin de résumer les deux phases de la photosynthèse.
On doit donc y retrouver tous les acteurs suivants : Complexe protéine-pigments photosynthétiques (ou
photosystème), ATPsynthétase, CO2, C5P2 (ou ribulose 1-5diphosphate), le C3P (ou triose phosphate), le PGA (ou
phosphoglycérate), autres sucres, amidon, acides aminés, acides gras. On indiquera bien les 3 étapes essentielles du
cycle de Calvin (Fixation du CO2, Réduction et Régénération)
1
/
4
100%