B10 - Les mécanismes de l`évolution

B10 - Les mécanismes de l’évolution
Connaissances de premier ordre indispensables
Capacités exigibles
1. Approche expérimentale de l’évolution
L’évolution correspond à un ensemble de modifications génotypiques d’une population. Ses mécanismes peuvent être approchés de façon expérimentale. Le
test de fluctuation de Luria & Delbrück a démontré, sur des bactéries, le caractère aléatoire des mutations. L’étude quantitative des populations de phalène
du bouleau en Grande-Bretagne a montré qu’un caractère (le mélanisme) favorisant la survie et la reproduction, dans un environnement industriel, voit sa
fréquence s'accroître d'une génération à l'autre. Dans ce cas, c’est essentiellement la prédation qui est le mécanisme de la sélection. Le caractère sélectionné
confère une adaptation au milieu dans lequel il est sélectionné.
L’expérience de Buri sur des populations expérimentales de drosophiles, non soumises à la sélection, montre qu’il se produit aussi des fluctuations aléatoires
des fréquences alléliques sous l’effet de la dérive génétique.
Figure B10.1
Montrer le caractère aléatoire des mutations (expérience de Luria &
Delbrück)
Définir les notions de sélection et d’adaptation (mélanisme de la
Phalène du bouleau, Figure B10.2) et de dérive (expérience de Buri,
Figure B10.3).
2. La sélection, un processus de reproduction différentielle
Chacune des caractéristiques biologiques, écologiques, comportementales d’un organisme qui est le résultat de l’évolution constitue un trait d’histoire de
vie. La sélection ne peut s’opérer que s’il existe au sein d’une population des traits héritables qui augmentent la valeur sélective de ceux qui les portent. . La
valeur sélective (« fitness ») d’un trait (allèle, génotype ou phénotype) se mesure au nombre de descendants produits par les individus porteurs de ce trait ;
elle est le produit de la viabilité (probabilité d’atteindre l’âge reproducteur) par la fertilité (nombre moyen de descendants viables). La valeur sélective d’un
trait dépend de plusieurs paramètres : pression de prédation, fréquence du trait dans la population, facteurs environnementaux, dynamique spatio-
temporelle,…
La sélection ne génère pas de diversité génétique ; elle exerce un tri orienté de la diversité génétique. Dans un environnement donné, la sélection
directionnelle modifie la moyenne et diminue la variance du caractère héritable sélectionné, ce qui conduit à une modification des phénotypes. Si l’avantage
sélectif est dépendant de la fréquence du trait, ou si des pressions négative et positive se combinent, il se produit une sélection balancée qui peut entretenir
un polymorphisme.
Une adaptation peut être définie comme un trait nouveau, apparu chez un organisme et maintenu chez ses descendants par la sélection naturelle.
Figure B10.4
Montrer que la valeur sélective d’un trait génétique dépend de
l’environnement.
Figure B10.5
Différencier les notions de sélection directionnelle (cas de la Phalène du
Bouleau) et de sélection balancée (cas des proportions de mâles et de
femelles, §IIB2b).
3. La dérive génétique, un processus évolutif aléatoire
La dérive résulte du tri aléatoire des gamètes à chaque génération, ce qui fait varier les fréquences alléliques. Sous l’effet de la dérive, un allèle peut être soit
fixé soit perdu. Si la dérive génétique est la seule force évolutive à agir sur une population, le polymorphisme à un locus donné ne peut être que transitoire ;
cependant, à l’échelle de l’ensemble des populations de cette espèce, le polymorphisme sera globalement maintenu. La dérive agit de la même façon sur les
allèles neutres ou sous sélection, mais elle est la seule à agir sur les allèles neutres.
L’effectif efficace est l’effectif des individus participant réellement à la reproduction. C’est l’effectif d’une population théorique, (effectif constant, individus
hermaphrodites dont les croisements s’effectuent au hasard et simultanément, absence de sélection) pour laquelle la dérive s’effectuerait à taux constant.
La dérive conduit à une différenciation génétique progressive entre chaque population d’une espèce et donc à l’émergence de structures génétiques propres
à chacune d’entre elles. Ainsi, les populations isolées géographiquement, présentent un polymorphisme propre, représentatif du pool de gènes porté par les
premiers migrants : c’est l’effet de fondation qui joue un rôle de tout premier plan dans la formation de nouvelles espèces. À l’échelle des temps
géologiques, la perte de diversité des dinosaures grands herbivores à la fin du Crétacé, a contribué à la disparition des dinosaures non aviens, par un effet de
goulot d’étranglement, libérant alors des niches écologiques, ce qui a favorisé l’augmentation du taux de spéciation chez les mammifères au début du
Tertiaire.
§IIIA1. Expliquer l’action de la dérive sur les traits neutres et
sélectionnés.
§IIIB2. Savoir définir l’effectif efficace.
Aucun calcul n’est requis.
Présenter deux exemples de dérive, à deux échelles d’étude :
dérive génétique au sein d’une population : cas de l’effet fondateur
sur les fréquences alléliques §IIIC1;
perte de diversité des dinosaures lors de la crise KT remplacés par les
Mammifères dans des niches écologiques comparables (constat à
réaliser sur la niche des grands herbivores) - dérive
phylogénétique§IIIC2.
4. Isolement génétique et spéciation
Chez les Eucaryotes, une espèce biologique est « un groupe de populations naturelles actuellement interfécondes et reproductivement isolées des autres
groupes équivalents ». Les définitions de l’espèce peuvent aussi intégrer d’autres critères : phénotypique (ressemblance), écologique (partage d’une niche
écologique), phylogénétique (partage d’une relation d’ascendance et de descendance). Les limites géographiques de l’espèce constituent son aire de
distribution.
L’isolement reproducteur est le moteur de la spéciation. Les processus d’isolement empêchent la formation des zygotes hybrides (isolement pré-zygotique)
ou réduisent leur valeur sélective (isolement post-zygotique). Néanmoins, les transferts horizontaux et les hybridations sont des limites à ces isolements,
même si l’hybridation peut aussi participer au renforcement de l’isolement.
Les espèces ne sont pas pérennes : elles se forment par spéciation, et disparaissent. On distingue deux grands modèles de spéciation, suivant les modalités
géographiques de l’apparition d’une divergence entre deux populations. Dans le modèle de spéciation allopatrique, le flux génique entre deux populations
est interrompu par une barrière géographique. Dans le cas d’une spéciation sympatrique, la divergence entre deux sous-populations s’acquiert sans barrière
géographique entre elles ; elle peut être la conséquence d’une allopolyploïdisation (cas des Spartines européennes). L’hybridation interspécifique, quand
elle est possible, peut être à l’origine d’espèces nouvelles (évolution réticulée)
L’évolution n’a ni direction, ni but ; elle ne peut donc pas être présentée en termes de «progrès» et elle peut être simplificatrice. Tous les organismes
évoluent : en ce sens, il n'y a ni fossile vivant, ni organisme primitif.
§IVA. Présenter les différents critères susceptibles de fonder d’autres
définitions de l’espèce (phénotypique, écologique, phylogénétique) ;
§IVB1 à 3. Discuter, pour les Eucaryotes, la notion d’hybridation dans le
contexte de l’espèce biologique ;
§IVB4. Discuter la notion d’espèce chez les procaryotes en lien avec les
transferts génétiques horizontaux ;
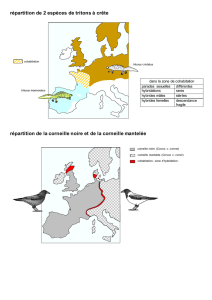
§IVC Manipuler deux exemples de spéciation (un exemple sympatrique,
les Spartines européennes (figures B10.10 et 11) et un exemple
allopatrique (figure B10.9) ;
Présenter la notion d’évolution réticulée (à l’aide des deux points
précédents : hybridation et transferts horizontaux).

1
/
2
100%