LE CONTRÔLE MOTEUR-13

1

La posture est un arrangement des segments corporels qui reflète la géométrie
segmentaire à un temps donné.
Chez l’homme 60% de la musculature est dédiée à la résistance à la gravité.
Tout mouvement est accompagné d’ajustements posturaux qui visent au maintien
de l’équilibre.
Des ajustements posturaux anticipateurs apparaissent quelques millisecondes
avant la réalisation d’un mouvement volontaire.
2

3

4
Les muscles squelettiques assurent le mouvement des os autour des articulations. On distingue :
les muscles axiaux qui commandent les mouvements du tronc (posture)
Les muscles proximaux qui commandent la partie proximale des membres (locomotion)
Les muscles distaux qui commandent les mouvement des articulations des mains, des pieds et des
doigts (mouvements fins)
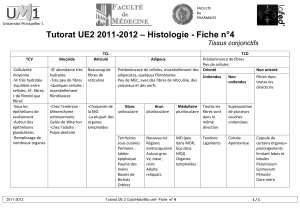
Chaque muscle est emballé dans un tissu conjonctif dont les extrémités forment les tendons qui
s’attachent aux articulations.
Un muscle est formé de plusieurs centaines de fibres musculaires.
Il existe plusieurs types de fibres, essentiellement :
- les fibres rapides (type II), pauvres en mitochondries, se contractent fortement et rapidement
mais se fatiguent rapidement (métabolisme anaérobie);
- les fibres lentes (type I), riches en mitochondries ont des vitesses de contraction lentes mais
soutiennent une contraction continue sans fatigue (métabolisme aérobie).
Chaque muscle est composé de fibres rapides ou lentes, en proportion différente suivant sa
fonction : muscles rapides (blancs) ou lents (rouges).
Les contractions des fibres musculaires sont déclenchées par des motoneurones α
αα
α, par
l’intermédiaire d’une synapse utilisant l’acétylcholine comme médiateur. Les corps des
motoneurones α
αα
αsont situés dans les cornes ventrales de la moelle épinière.
Chaque fibre ne reçoit qu’un moto neurone α
αα
α, mais un motoneurone α
αα
αpeut commander un grand
nombre de fibres (plus de mille par exemple dans les muscles antigravitaires de la jambe). Un
motoneurone α
αα
αn’innerve qu’un type de fibres. Un motoneurone + l’ensemble des fibres qu’il
innerve constitue l’unité motrice.
Les corps cellulaires des motoneurones sont regroupés en amas au niveau de la moelle épinière,
où ils forment des noyaux moteurs.
Le contrôle de la contraction musculaire se fait par la fréquence de décharge des unités motrices,

et par le recrutement de nouvelles unités motrices.
4
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
1
/
36
100%