H P T 16

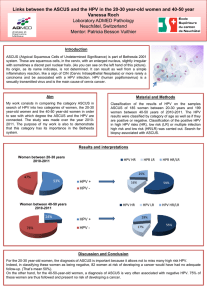
HUMAN PAPILLOMAVIRUS TYPE 16
IN HEAD AND NECK CARCINOGENESIS
REN WEI CHEN
HELSINKI 2004

Helsinki University Biomedical Dissertations No. 54
Human Papillomavirus Type 16
in Head and Neck Carcinogenesis
Ren Wei Chen
Department of Virology
Haartman Institute
and
Department of Otorhinolaryngology −
Head and Neck Surgery
Helsinki University Central Hospital
University of Helsinki
Finland
Academic Dissertation
To be publicly discussed, with the permission of the Faculty of
Medicine of the University of Helsinki, in the Small Lecture Hall,
Haartman Institute, on June 18th at 12 o’clock noon.
Helsinki, 2004

Human papillomavirus type 16 in head and neck carcinogenesis
2
Supervised by: Professor Antti Vaheri
Department of Virology
Haartman Institute
University of Helsinki
Helsinki, Finland
and
Doctor Leena-Maija Aaltonen
Department of Otorhinolaryngology
<
Head and Neck Surgery
Helsinki University Central Hospital
Helsinki, Finland
Reviewed by:
Department of Oral Pathology and Radiology
Institute of Dentistry
University of Turku
Turku, Finland
and
Professor Reidar Grenman
Department of Otorhinolaryngology
<
Head and Neck Surgery
Turku University Central Hospital
Turku, Finland
Opponent: Doctor Peter J.F. Snijders
Department of Pathology
Vrije Universiteit Medical Center
Amsterdam, The Netherlands
ISSN 1457-8433
ISBN 952-10-1911-5 (Paperback)
ISBN 952-10-1912-3 (PDF)
http://ethesis.helsinki.fi
Edita Prima Oy
Helsinki, 2004
Professor Stina Syrjänen

Table of contents 3
TABLE OF CONTENTS
1. ABSTRACT ........................................................................................................... 5
2. ORIGINAL PUBLICATIONS.................................................................................. 7
3. ABBREVIATIONS AND DEFINITION OF SOME ESSENTIAL TERMS................ 8
4. INTRODUCTION.................................................................................................. 10
5. REVIEW OF THE LITERATURE ......................................................................... 12
5.1. HUMAN PAPILLOMAVIRUS ......................................................................... 12
5.1.1. Classification of human papillomaviruses ............................................... 13
5.1.2. HPV life cycle .......................................................................................... 14
5.1.3. Function of viral proteins ......................................................................... 14
5.2. HISTORICAL BACKGROUND OF HPV AND CANCER............................... 18
5.3. PTEN AND LKB1 TUMOR SUPPRESSOR GENES ..................................... 18
5.4. SQUAMOUS CELL CARCINOMA OF THE HEAD AND NECK..................... 19
5.4.1. Etiology and risk factors for the head and neck cancer........................... 20
5.4.2. Tumor classification ................................................................................ 21
5.4.3. HPV in HNSCC ....................................................................................... 23
5.4.4. HPV in SCC of tonsils ............................................................................. 25
5.4.5. HPV DNA in tumor-free tonsils................................................................ 25
6. AIMS OF THE STUDY ......................................................................................... 27
7. PATIENTS, MATERIALS, AND METHODS ........................................................ 28
7.1. PATIENTS AND MATERIALS (I, II, III) .......................................................... 28
7.2. CELL LINES (IV)............................................................................................ 28
7.3. PLASMIDS (IV).............................................................................................. 29
7.4. METHODS..................................................................................................... 29
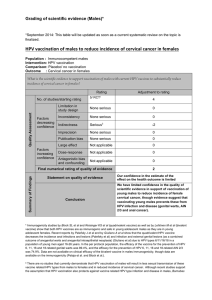
8. RESULTS AND DISCUSSION ............................................................................ 30
8.1. SOMATIC MUTATION IN PTEN AND LKB1 GENES IS A RARE EVENT IN
LARYNGEAL TUMORS (I) ................................................................................... 30
8.2. PREVALENCE AND PHYSICAL STATUS OF HPV IN HNSCC (II) .............. 31
8.2.1. Prevalence, genotype, viral load, and physical status of HPV DNA in
HNSCC ............................................................................................................. 31
8.2.2. Absence of viral capsid protein expression in HNSCC............................ 33
8.2.3. Correlation of HPV DNA status and clinical outcome.............................. 34

Human papillomavirus type 16 in head and neck carcinogenesis
4
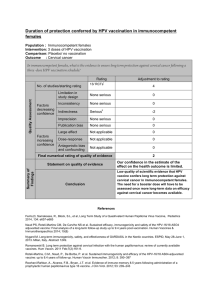
8.3. PRESENCE OF HPV-16 DNA BUT NO OTHER TYPES IN TUMOR-FREE
TONSILS (III)........................................................................................................ 35
8.3.1. Presence of HPV DNA and HPV genotypes in tumor-free tonsils........... 35
8.3.2. Seroprevalence of antibodies against HPV-16 L1, E6, and E7 ............... 36
8.3.3. Non-concordance between presence of HPV DNA in tonsillar tissue and
seropositivity to HPV proteins ........................................................................... 37
8.4. ESTABLISHMENT AND CHARACTERIZATION OF HPV-16 DNA
IMMORTALIZED HUMAN TONSILLAR EPITHELIAL CELL LINES (IV) .............. 37
8.4.1. Growth and differentiation of HPV-16 DNA transfected human tonsillar
epithelial cells.................................................................................................... 37
8.4.2. Tumorigenicity of HPV-16 transformed cell lines..................................... 38
8.4.3. Cytogenetic aberration ............................................................................ 39
8.4.4. Physical status and chromosomal localization of viral DNA in HPV-16
transformed HTE cell lines ................................................................................ 40
8.4.5. Viral gene expression in HPV-16 transformed HTE cell lines.................. 41
9. CONCLUDING REMARKS.................................................................................. 42
10. PERSPECTIVES................................................................................................ 43
11. ACKNOWLEDGEMENTS.................................................................................. 44
12. REFERENCES................................................................................................... 46
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
1
/
67
100%