annexe - Catalogue en ligne La Bibliothèque Universitaire Centrale

REPUBLIQUE ALGERIENNE DEMOCRATIQUE ET POPULAIRE
MINISTERE DE L’ENSEIGNEMENT SUPERIEUR ET DE LA RECHERCHE SCIENTIFIQUE
N°d’ordre ……..
Série………….
Mémoire
Présenté pour l’obtention du diplôme :
De magister en médecine vétérinaire
Option : pathologie
Spécialité aviculture et pathologie aviaire
Par Meme CHOUDER. Nedjma .N ée BELKACEMI
SOUTENU LE ...08../…07../2006
Jury de soutenance
Président ELHADEF ELOUKKI S Professeur Université de Constantine
Rapporteur BENMAKHLOUF A Maître de conférences
Université de Constantine
Examinateur AOUN L Maître de conférences
Centre El Tarf
MAMMACHE B Maître de conférences
Université de Batna
2005-2006
Université Mentouri Constantine
Faculté des sciences
Département des sciences vétérinaires

Dédicaces
A mon père,
Q’il puisse reposer en paix
A ma mère,
Ma douce et tendre mère, qui a tant sacrifié pour notre réussite, q’elle puisse trouver
dans mon travail, le fruit de son labeur.
A mes sœurs,
La gentille Hayet,
La très sage Amel,
La généreuse, au cœur d’or, Nadia,
L’ange, Nabila et la très douce Samia ;
Je vous remercie toute de m’avoir encouragé à continuer mon Magister et soutenu
dans les moments les plus délicats.
Tout particulièrement, à toi Abdel Aziz,
Mon cher et unique frère, soit fier de moi aujourd’hui
A mon cher et tendre époux Kamel Eddine,
Je n’oublierais jamais ta patience avec moi et ta grande générosité.
A, Yahia El Hakim,
Mon adorable petit bonhomme, qu’il soit fier de sa maman
A, mes beaux parents,
Vers les quels j’ai un grand respect, je vous remercie de vous occuper avec
amour et passion de mon fils Yahia.
A, ma belle sœur Ilhem,
A qui je souhaite une bonne adaptation à l’étranger, et surtout une réussite dans
Ses études.
A, mes beaux frères,
Que Dieu tout puissant vous préserve du mal et vous aide à réussir votre vie
professionnelle.
A, touts mes collègues,
La grande dame Chérifa,
La belle et glorieuse Nadjoua,
La sympathique Meryam,
L’honorable Abdel Wahab,
Le rigoureux Amir, le gentil Hmed, et tout particulièrement le très généreux
Saber,
A, la fleur de la promotion,
A toi Nawel, je te souhaite de réussir brillamment tes études ainsi que ta vie
privé.
Je vous souhaite à tous une bonne réussite.

Dédicaces
A la mémoire de monsieur O. Hadad,
Q’il repose en paix, et que Dieu tout puissant puisse l’accueillir dans
son vaste Paradis.
A mon encadreur, monsieur A. Benmakhloouf,
Je vous remercie de nous avoir poussé dans la bonne voie, celle du
travail et de la patience, et consacré un peu de votre temps précieux
pour corriger nos thèses.
Au Directeur du laboratoire régional vétérinaire, monsieur A. Boukerou,
Je vous remercie du fond du cœur, de m’avoir permis de travailler au
sein de votre laboratoire, et m’avoir fait confiance pendant toute la
durée du travail.
A monsieur Ben Segni,
Que je respecte profondément, je n’oublierais jamais lorsqu’il nous a
dit : « il n’y a pas de travail, sans résultats ». Cette phrase m’a aidé à
avoir confiance en moi et continuer mon travail jusqu'à aboutissement.
A tout le personnel du laboratoire régional,
Qui ont montré une grande gentillesse et sympathie en vers moi, tout
particulièrement, Salima du service de la bactériologie alimentaire,
Noujoud du service de virologie et mon amie Roufia.
Plus spécialement, tout le personnel du service de bactériologie médicale,
Qui m’ont accueilli chaleureusement et mon laissé travaillé en toute
facilité.
N. Chouder.

Remerciement,
Mes profonds remerciements sont adressés au président du jury,
Notre respectable professeur, monsieur ELHADEUF EL OKKI S, qui a accepté de
présider le jury de soutenance.
A, Madame AOUN L, maître de conférence au centre EL Tarf
Et monsieur MAMACHE B, maître de conférence à l’université de Batna
A monsieur Benmakhloouf A, maître de conférence à l’université de Constantine
Je tiens à vous remercier de m’honorer par votre présence en temps qu’examinateurs et
encadreur.
Je vous remercie tous d’apporter votre savoir et sagesse en jugeant mon
modeste travail
Chouder N

SOMMAIRE
Titre Page
INTRODUCTION
RAPPELS ANATOMIQUES DE L’APPAREIL DIGESTIF DES
OISEAUX
1. L’APPAREIL DIGESTIF ET SES ANNEXES………………………
1.1. DESCRIPTION……………………………………………………….
1.1.1. LA REGION CRANIALE DU TUBE DIGESTIF…………………
1.1.2. LA REGION STOMACALE DU TUBE DIGESTIF………………
1.1.3. LA REGION POSTERIEURE DU TUBE DIGESTIF……………..
1.2. ANATOMIE DE LA REGION POSTERIEURE DU TUBE
DIGESTIF …………………………………………………………………
1. 2.1 LE DUODENUM ………………………………………………..
1.2.2 LE JEJUNUM ET L’ILEON OU TRACTUS DE
MECKEL………………………………………………………………….
1.2.3 LE RECTUM………………………………………………………
1.2.4 LES CAECUMS……………………………………………………
1.2.5 LE CLOAQUE……………………………………………………..
1.3 STRUCTURES CYTOLOGIQUES DE LA PAROI DU TUBE
DIGESTIF.....................................................................................................
1.3.1 LES ENTEROCYTES……………………………………………………
1.3.2 LES CELLULES…………………………………………………………
1.3.3 LES ENZYMES……………………………………………………………
1.3.4 LES MICELLES……………………………………………………………
2. DEVELOPPEMENT DI TRACTUS GASTRO-INTESTINAl…….
2.1 DEVELOPPEMENT GENERAL ……………………………………
2.2 DEVELOPPEMENT AU STADE EMBRYONNAIRE ……………..
2.3 DEVELOPPEMENT CHEZ L’ADULTE…………………………….
RAPPELS BACTERIOLOGIQUES
1. IDENTIFICATION BACTERIOLOGIQUE DE QUELQUES
ESPECES DE LA FAMILLE DES
ENTEROBACTERIACEAE .....................................................................
..
1.1 MORPHOLOGIE ET CLASSIFICATION…………………………
1.1.1 DEFINITION ET IMPORTANCE………………………………..
1.1.2. CARACTERES CULTURAUX…………………………………
1.2 POUVOIR PATHOGENE…………………………………………
1.2.1 NOTION DE POUVOIR PATHOGENE ………………………………
..……………01
..……………01
…………….01
……………..01
……………..02
……………..02
…………….02
……………..03
…………….03
…………….03
…………….04
………….….04
….…………04
……...………05
………………07
…………….07
…………….08
………………08
…………….09
…………..10
……………11
………………11
..……………..11 ………………12
……………12
……………12
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
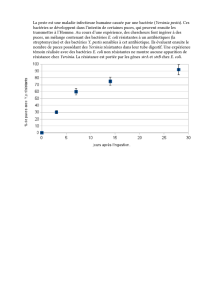
1
/
190
100%