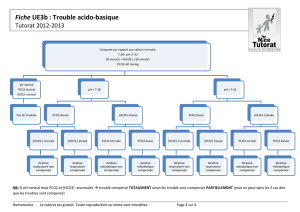
L`appareil respiratoire et les échanges gazeux

1
1ère Année.
L’appareil respiratoire et les échanges gazeux
2004
J.P. PERRET

2
L’appareil respiratoire et les échanges gazeux
1. Anatomie de l’appareil respiratoire : p. 4
1.1 Les voies aériennes :
- trachée, bronches , bronchioles, épithélium cylindrique cilié
- alvéoles, membrane respiratoire, épithélium pavimenteux simple
- le surfactant alvéolaire
1.2 Les poumons et la plèvre
1.3 Les muscles respiratoires.
1.4 L’irrigation sanguine pulmonaire et systémique
1.5 La bronchomotricité
2. Les échanges gazeux p. 6
2.1 Modalité : la diffusion
2.2 La pression partielle de l’O2 et du CO2 :
- dans l’air atmosphérique
- dans l’air alvéolaire et le sang veineux pulmonaire
- dans les tissus et le sang veineux systémique
2.3 Le transport des gaz dans le sang : p. 7
2.3.1 Oxygène :
- dissous
- combiné :- courbes de (dé)saturation de l’hémoglobine : influence de la
température, de la pCO2 et du DPG
- courbe de (dé)saturation de la myoglobine
2.3.2 Dioxyde de carbone : p. 8
- dissous
- carbaminé
- sous forme de bicarbonates : le rôle des hématies
3. La régulation de la ventilation : (mouvements des muscles respiratoires)
3.1 Les volumes pulmonaires p. 9
3.2 L’organisation du système de contrôle volontaire et automatique de la ventilation.
3.3 Les réponses ventilatoires et leurs stimuli d’origine : p. 11
4. Les système tampons du sang et le rôle de la ventilation dans la p. 12
régulation de l’équilibre acido-basique : ( voir aussi : le rôle du rein )
4.1 Le tampon plasmatique ac.carbonique- bicarbonate p. 13
La relation d’Henderson-Hasselbach
4.2 Le tampon intracellulaire oxyhémoglobine- désoxyhémoglobine réduite p. 14
4.3 La réponse ventilatoire ( et rénale ) aux variations du pH plasmatique :
4.3.1 : réponse à l’acidose
4.3.2 : réponse à l’alcalose et à la diminution de pO2
Testons nos connaissances : p. 16

3
Ce qu’il faut connaître et savoir
Connaître :
- la structure de la membrane respiratoire
- le rôle du surfactant alvéolaire
- le rôle de la plèvre
- le rôle des divers muscles respiratoires
- l’irrigation sanguine des poumons
- l’innervation des muscles respiratoires et des muscles lisses des bronches
- l’équation d’Henderson-Hasselbach
Savoir :
- expliquer comment s’effectuent les échanges de gaz respiratoires au niveau des
capillaires sanguins alvéolaires et tissulaires
- expliquer les échanges d’O2 à partir des courbes de (dé)saturation de
l’hémoglobine et de la myoglobine
- expliquer le transports de CO2 sous forme de bicarbonates
- expliquer à partir de la relation d’Henderson-Hasselbach comment les
modifications de la pCO2 et de la concentration des anions bicarbonates
influencent la ventilation
( et l’élimination rénale des bicarbonates : cf. chapitre « le rein » )

4
L’appareil respiratoire et les échanges gazeux
1 Anatomie de l’appareil respiratoire :
1.1 Les voies aériennes :
La paroi de la trachée et de ses ramifications, bronches et bronchioles, (fig. 1 ) est
maintenue ouverte par des anneaux semi circulaires de tissus cartilagineux, réunis par du
tissus conjonctif fibreux et des fibres musculaires lisses dont la contraction règle le
diamètre des bronches et bronchioles ( = bronchomotricité ). La face interne de cette
paroi est recouverte par un épithélium de type cylindrique simple, cilié, qui élabore du
mucus piégeant les microparticules ayant pénétré dans l’arbre respiratoire et les refoulant
jusqu’à l’entrée de la trachée.
L’extrémité des bronchioles est renflée en alvéoles groupées en sacs alvéolaires
La paroi des alvéoles n’est formée que par l’épithélium qui, à ce niveau est de type
pavimenteux simple non cilié. C’est là uniquement qu’ont lieu les échanges gazeux entre
l’air alvéolaire et le sang, à travers une structure appelée membrane respiratoire.
La membrane respiratoire ( fig. 2 ) est constitué par l’épithélium alvéolaire,
l’endothélium capillaire et leurs membranes basales. L’épaisseur de l’ensemble est de 0,5
µm.
L’air alvéolaire renferme une proportion élevée de vapeur d’eau qui tapisse la paroi
alvéolaire. Or la tension superficielle de l’eau devrait conduire à en rassembler les
molécules en une gouttelette qui provoquerait l’aplatissement de l’alvéole et y
empêcherait l’entrée de l’air. Ceci n’a pas lieu parce que l’épithélium alvéolaire sécrète du
surfactant alvéolaire, molécule constituée par l’association d’un phospholipide et d’une
protéine, qui abaisse la tension superficielle de l’eau en formant une couche
monomoléculaire à la surface du film d’eau qui recouvre la paroi alvéolaire. La sécrétion
de ce surfactant est stimulée lors des inspirations profondes qui distendent la paroi
alvéolaires.

5
1.2 Les poumons et la plèvre ( cf. schéma de cours )
Chaque poumon est enveloppé dans un double feuillet de tissus conjonctif qui
constitue la plèvre. Le feuillet externe ou pariétal est accolé aux muscles de la paroi
thoracique et au diaphragme et le feuillet interne ou viscéral à la surface du poumon Ces
feuillets sont séparés par une faible épaisseur de liquide pleural, en dépression ( 4 mm Hg
) par rapport à l’air pulmonaire, qui permet leur glissement mais s’oppose à leur
séparation et assure donc le contact permanent entre la surface des poumons et la paroi
thoracique .
1.2 Les muscles respiratoires ( ventilatoires )
Sont des muscles squelettiques dont l’activité permet de modifier le volume de la
cage thoracique et donc le volume d’air contenu dans les voies aériennes ainsi que la
vitesse de circulation de l’air dans les bronches.
L’inspiration est assurée par la contraction des muscles intercostaux externe et
surtout du diaphragme ( innervé par le nerf phrénique ). L’expiration, passive sauf lors
d’un exercice, correspond au relâchement de ces muscles. L’expiration forcée observée
lors d’un exercice est assurée par la contraction des muscles intercostaux internes et des
muscles abdominaux qui appuient alors sur le diaphragme.
1.3 L’irrigation sanguine pulmonaire et systémique
Les capillaires de la circulation pulmonaire sont disposés autour des alvéoles pour
assurer les échanges gazeux pulmonaires. Le sang efférent rejoint la veine pulmonaire et
l’oreillette gauche ;
Le reste des voies aériennes est irrigué par les artères bronchiques issues de l’aorte.
Le sang efférent rejoint la veine cave supérieure
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
6
7
8
9
10
11
12
13
14
15
16
17
18
19
1
/
19
100%