Shigella DES - Pôle de Biologie Pathologie Génétique

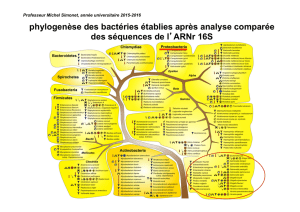
phylogenèse des bactéries établies après analyse comparée
des séquences de l’ARNr 16S (horloge évolutionnaire)
Professeur Michel Simonet, année universitaire 2013-2014

entérobactéries
arbre établi à partir des séquences du facteur d’élongation Tu
(International Journal of Systematic & Evolutionary Microbiology, 2005)
! des bacilles aérobies-anaérobies
! sans exigence culturale particulière
! cytochrome oxydase -
! catalase +
! nitrate-réductase +
! fermentant le glucose
γ-proteobactéries

Klebsiella pneumoniae
des diplobacilles entourés d’une capsule
Escherichia coli
des bacilles polymorphes à coloration bipolaire
des bactéries à Gram négatif

Klebsiella pneumoniae
Escherichia coli Proteus mirabilis
Serratia marcescens

Enterobacter cloacae
Klebsiella pneumoniae
Serratia marcescens
VP+
+
-
+ - - - -- + + +
+ -
+ -- - -- + + +
+
-
+ -- - - + + + +
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
1
/
35
100%



![Télécharger la fiche technique [120.43 Ko PDF]](http://s1.studylibfr.com/store/data/000674177_1-17ee923c1234f9f2353ebcf5ad682c82-300x300.png)






