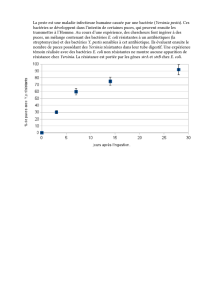
T H È S

.
T
TH
HÈ
ÈS
SE
E
En vue de l'obtention du
D
DO
OC
CT
TO
OR
RA
AT
T
D
DE
E
L
L’
’U
UN
NI
IV
VE
ER
RS
SI
IT
TÉ
É
D
DE
E
T
TO
OU
UL
LO
OU
US
SE
E
Délivré par L’Université Toulouse III – Paul Sabatier
Discipline ou spécialité : Pharmacologie
JURY
Monsieur le Professeur Alain BOUSQUET-MELOU Examinateur
Monsieur le Professeur Pascal GUSTIN Rapporteur
Monsieur le Professeur Georges HOUIN Examinateur
Madame le docteur Isabelle KEMPF Rapporteur
Monsieur le Docteur Michel LAURENTIE Examinateur
Monsieur le Professeur Pierre-Louis TOUTAIN Examinateur
Membre invité : Madame Agnès PERRIN-GUYOMARD
Ecole doctorale : Biologie-Santé-Biotechnologies
Unité de recherche : UMR 181 Physiopathologie et Toxicologique Expérimentales, INRA, ENVT
Laboratoire d’Etudes et de Recherches sur les Médicaments Vétérinaires et les Désinfectants, AFSSA
Directeur(s) de Thèse : Monsieur le Professeur Pierre-Louis TOUTAIN
Monsieur le Docteur Michel LAURENTIE
Monsieur le Professeur Alain BOUSQUET-MELOU
Rapporteurs : Monsieur le Professeur Pascal GUSTIN
Madame le docteur Isabelle KEMPF
Présentée et soutenue par Anne-Sylvie KESTEMAN
Le 14 décembre 2009
Titre : Influence des facteurs associés à une antibiothérapie de type métaphylactique sur les relations
pharmacocinétiques/pharmacodynamiques (PK/PD) des antibiotiques.
Conséquences sur les schémas posologiques et sur l’émergence de résistance


1
A Madame le Docteur Isabelle KEMPF, et Monsieur le Professeur Pascal GUSTIN, qui ont
accepté de consacrer de leur temps à la lecture et à l’appréciation de ce manuscrit, qu’ils
trouvent ici le témoignage de notre respectueuse gratitude.
A Monsieur le Professeur Georges HOUIN, qui nous a fait l’honneur d’accepter de faire partie de
notre jury de thèse. Qu’il soit assuré de nos plus sincères remerciements.
A Messieurs les Professeurs Alain BOUSQUET-MELOU et Pierre-Louis TOUTAIN, qui nous ont
accueillie comme doctorante, nous ont aidés et guidés tout au long de ces années de thèse avec
rigueur et optimisme. Qu’ils soient assurés de notre sincère reconnaissance.
A Messieurs les Docteurs Michel LAURENTIE et Pascal SANDERS, qui nous ont accueillie au sein
de leur laboratoire. Qu’ils soient assurés de tout notre respect et de toute notre reconnaissance.
A tous les membres de l’équipe « Antibiothérapie et Antibiorésistance » de l’UMR 181 qui ont
contribué à la réalisation de ce travail : Madame Nathalie ARPAILLANGE, Madame le Docteur
Véronique DUPOUY, et Madame Sylvie PUEL. Sincères remerciements.
A toute l’équipe zootechnie de l’UMR181 : Monsieur Sylvain BRUYAS, Monsieur Jean-Pierre GAU,
Monsieur Joseph MALIGOY, Monsieur Patrice ROUBY et Monsieur René SCHAN. Un grand merci
pour leur aide précieuse.
A tous les membres de l’unité Pharmacocinétique Pharmacodynamique du LERMVD de l’AFSSA de
Fougères qui m’ont accueillis chaleureusement et ont contribué à la réalisation de ce travail :
Madame Annick Brault, Madame le Docteur Mireille BRUNEAU, Mademoiselle Pamela LOUÂPRE,
pour sa bonne humeur et sa disponibilité, Madame Jacqueline MANCEAU, et Madame Catherine
POIRIER. Sincères remerciements.

2
A Madame Agnès PERRIN-GUYOMARD, pour ses conseils, son soutien, ses critiques toujours
constructives, pour s’être investie dans ce travail de fourmi, en souvenir de toutes ces boîtes
ensemencées et comptées un très grand merci.
A l’équipe zootechnie du LERMVD, Monsieur Stéphane MARTEAU, Monsieur Jean-Guy ROLLAND,
mes deux anges gardiens de Fougères. Un grand merci pour leur précieuse aide durant les phases
animales, pour leur inventivité, pour leur bonne humeur et leur soutien durant les longues
journées d’expérimentation.
A toute l’équipe des thésards, anciens et actuels à Toulouse : Julie ANTIC, Delphine BIBBAL
pour sa bonne humeur et toutes nos discussions, Séverine COLLET pour son aide précieuse sur la
dernière ligne droite, Aude FERRAN ma binôme, pour tous ses conseils, son immense patience et
ces heures passées enfermées dans notre bocal !, Elizabeth JEUNESSE, Julien LEGHAIT, pour
son initiation à la pose de cath, son calme et son optimisme et Béatrice ROCQUES, bon courage à
elle qui débute. Merci à tous pour votre bonne humeur et les tous les bons moments passés
ensemble.
A Fougères : Emmanuel BAZIN, Julie DAVID, merci de m’avoir initiée aux joies du pays
fougerais, pour nos ballades, tous ces moments où l’on a refait le monde, et pour m’avoir accueilli
chez elle de façon si spontanée !, Stéphanie FAURE, et Jérôme HENRI, merci à tous pour votre
accueil et tous ces bons moments passés ensemble.
A tous les membres de l’AFSSA Fougères, pour leur bonne humeur et leurs encouragements, avec
une mention spéciale aux « Cantine’s squatters » avec qui j’ai passé d’excellent moments
musicaux.
A tous les membres de l’UMR181, pour toute l’aide indispensable, qu’elle soit technique,
administrative ou scientifique qu’ils m’ont apportée, leur soutien, leur bonne humeur et leurs
encouragements. Un immense merci.
A ma famille pour leur amour, leur soutien et leurs encouragements tout au long de mes études.
A Emmanuel pour son amour, pour sa confiance et ses conseils

3
AVANT PROPOS
Ces travaux de thèse ont fait l’objet, à l’heure actuelle, des publications et communications
suivantes :
PUBLICATIONS
A.-S. KESTEMAN, A PERRIN-GUYOMARD, M. LAURENTIE, P. SANDERS, P.-L. TOUTAIN
and A. BOUSQUET-MELOU
Emergence of Resistant Klebsiella pneumoniae in the Intestinal Tract during
Successful Treatment of Klebsiella pneumoniae Lung Infection in Rats.
Antimicrobial Agents and Chemotherapy, 2010 Jul; 54(7):2960-64
A.-S. KESTEMAN, A.A. FERRAN, A PERRIN-GUYOMARD, M. LAURENTIE, P. SANDERS,
P.-L. TOUTAIN and A. BOUSQUET-MELOU
Influence of Inoculum Size and Marbofloxacin Plasma Exposure on the Amplification
of Resistant Subpopulations of Klebsiella pneumoniae in a Rat Lung Infection Model.
Antimicrobial Agents and Chemotherapy, 2009 Nov; 53(11):4740-48
A.A. FERRAN, A.-S. KESTEMAN, P.-L. TOUTAIN and A. BOUSQUET-MELOU
Pharmacokinetic/Pharmacodynamic Analysis of the Influence of Inoculum Size on the
Selection of Resistance in Escherichia coli by a Quinolone in a Mouse Thigh Bacterial
Infection Model.
Antimicrobial Agents Chemotherapy. 2009 Aug;53(8):3384-90
COMMUNICATIONS ORALES
A.-S. KESTEMAN, A.A. FERRAN, A PERRIN-GUYOMARD, M. LAURENTIE, P. SANDERS,
P.-L. TOUTAIN and A. BOUSQUET-MELOU
Influence of Inoculum size and Dosing Regimen on the Selection of Marbofloxacin
Resistant Mutants in Experimental Rat Klebsiella pneumoniae Lung Infection.
11th International Congress of the European Association for Veterinary Pharmacology and
Toxicology (EAVPT), 2009, Leipzig, Germany
A.-S. KESTEMAN, A PERRIN-GUYOMARD, M. LAURENTIE, P. SANDERS, P.-L. TOUTAIN
and A. BOUSQUET-MELOU
Impact d’une fluoroquinolone sur la flore pathogène et la flore commensale intestinale
dans un modèle d’infection pulmonaire par Klebsiella pneumoniae chez le rat
dixénique.
Journée d’Animation Scientifique du département santé animale de l’INRA, 2009, Port
d’Albret, France
A.-S. KESTEMAN, A.A. FERRAN, A PERRIN-GUYOMARD, M. LAURENTIE, P. SANDERS,
P.-L. TOUTAIN and A. BOUSQUET-MELOU
Influence of inoculum size and dosage regimens on the selection of marbofloxacin
resistant mutants in experimental rat Klebsiella pneumoniae lung infection.
4th International conference on Antimicrobial Agents in Veterinary Medicine (AAVM), 2008
Prague, République Tchèque
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
1
/
185
100%