9 — tapia

117
9—
MalaissePinelRazafiManantsoa
RakotoniainaRakotondRasoaRatsiMba
bogaeRtHaubRugeVeRHeggen

Les vers à soie malgaches
118

119
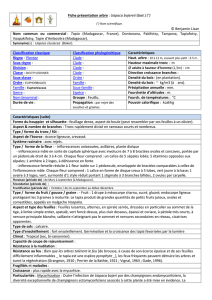
Aspects botaniques et écologiques des formations de tapia

Les vers à soie malgaches
120
Station Altitude
(m)
P
(mm)
T
(°C)
HR
(%)
N TM
(°C)
Tm
(°C)
Arivonimamo 1451 1460,1 17,8 79 121 26,1 7,8
Ambatonandrahana 1112 1288,8 18,3 75 104 30,8 7,7
Figure 9.1.Diagrammesombrothermiquesd'Arivonimamoetd'Ambatonandrahana
(d'aprèsDirectionGénéraledelaMétéorologie,Madagascar).

121
Aspects botaniques et écologiques des formations de tapia
 6
7
8
9
10
11
12
13
14
15
16
6
7
8
9
10
11
12
13
14
15
16
1
/
16
100%