LA PHYSiOlOGiE DE l`EXERCiCE

Physiologie*de*l’exercice*–*B.*BARON*et*Pr.*P.*PELAYO*
1*
1
La Physiologie de
l’exercice
Auteurs
Bertrand BARON, Université de la Réunion
Pr. Patrick PELAYO, Université Lille 2

2*
Physiologie*de*l’exercice*–*B.*BARON*et*Pr.*P.*PELAYO*
2
Chapitre 1 : Le muscle : transformateur d’énergie.
1. Introduction.
Toutes activités physiques et sportives impliquent une interaction entre le sportif et un
engin, le sol ou un autre sportif. Ainsi le lanceur va interagir avec un poids, un disque ou
un marteau, le coureur à pied avec le sol, alors que le judoka va interagir directement avec
un adversaire. Dans tous les cas, ces différentes interactions sont possibles dans un
premier temps grâces à des mouvements segmentaires, c'est-à-dire au déplacement d'un
segment osseux par rapport à un autre. Ainsi pour lancer le poids le plus loin possible, le
lanceur va effectuer une extension du bras sur l'avant-bras afin d'accélérer le poids.
L’action ne va pas se limiter à ce seul mouvement bien entendu, le tronc et les membres
inférieurs participeront également. En fait, même le plus simple mouvement localisé,
comme lorsqu’on soulève une charge en flexion de l’avant bras sur le bras, entraîne la
contraction de plusieurs muscles participant directement au mouvement (ce sont les
muscles dits agonistes : dans le cas de la flexion de l’avant bras sur le bras : biceps
brachial mais également le brachial et le long supinateur) mais aussi la contraction de
nombreux autres muscles participant au contrôle de la posture notamment.
Autres exemples : le coureur à pied va effectuer une extension du pied sur la jambe et de
la jambe sur la cuisse afin de se déplacer vers l'avant après contact avec le sol.
Ou encore, le judoka effectuera une flexion de l'avant-bras sur le bras afin d'attirer son
adversaire vers lui. On pourrait multiplier ces exemples à l'infini.
Tous ces mouvements segmentaires sont rendus possibles grâce à l'action des muscles qui
développent une force permettant de déplacer les leviers osseux autour des différents axes
articulaires (le coude, le genou, la cheville par exemple). Cette force est produite par la
transformation d'énergie chimique en énergie mécanique au sein du muscle. Et c'est grâce
aux 600 et quelques muscles que comporte notre organisme que l'on peut marcher, courir,
lancer, sauter, nager, frapper, danser, et faire tous les mouvements et activités physiques
possibles et imaginables.
2. Structure du muscle.
Le muscle est constitué de milliers de cellules musculaires de forme cylindrique qu'on
appelle les fibres musculaires. Plusieurs fibres musculaires forment un faisceau et
plusieurs faisceaux forment l'ensemble du muscle. Chaque fibre musculaire est entourée
par une fine couche de tissu conjonctif : l’endomysium qui la sépare des autres fibres. Le
sarcolemme, situé sous l’endomysium, délimite le contenu cellulaire. De même chaque
faisceau musculaire est entouré du périmysium, alors que l’épimysium entoure l'ensemble
du muscle. Ces différentes enveloppes conjonctives se réunissent pour former un tissu

Physiologie*de*l’exercice*–*B.*BARON*et*Pr.*P.*PELAYO*
3*
3
dense et très solide : le tendon. Elles se sont entrelacées avec les fibres de l’os. Cet
ensemble constitue donc un lien très solide entre le muscle et l’os.
Le muscle est composé d'environ 75 % d’eau. Mais les éléments les plus importants d’un
point de vue fonctionnel sont les protéines, qui correspondent à environ 20 % de la masse
musculaire. Si elles sont si importantes, c'est qu'elles sont responsables des propriétés
contractiles du muscle.
Les principales protéines sont la myosine, l’actine, la troponine et la tropomyosine.
Lorsqu'on observe une fibre musculaire au microscope, on peut observer une alternance
de bandes claires et de bandes sombres qui lui donnent un aspect strié. En langage
technique les bandes claires sont dites isotropes, ce qui signifie qu’elles laissent passer la
lumière. C'est pourquoi on les appelle les bandes I (I pour isotropes). Si elles laissent
passer la lumière, c'est parce qu'à cet endroit, seuls les filaments fins constitués
principalement d'actine et de tropomyosine sont présents.
A l'inverse les bandes sombres sont dites anisotropes et on les appelle bandes A. c'est
parce qu’à cet endroit on peut observer la présence des filaments fins mais également des
filaments épais constitués de myosine.
La bande I est séparée en deux parties par la ligne Z. L'unité comprise entre 2 lignes Z
s'appelle le sarcomère. C'est l'unité fonctionnelle de la cellule musculaire. C'est à
l'intérieur de cette unité fonctionnelle que la contraction musculaire va se dérouler,
principalement sous l’effet d’une interaction entre les filaments fins et épais aboutissant à
un raccourcissement du sarcomère.
3. Approche mécanique de la contraction musculaire.
Lors de la contraction, la force musculaire est transmise au segment osseux sur lequel le
muscle s’insère. Les muscles ayant un point d'insertion haut et bas sur 2 segments osseux
séparés par une ou plusieurs articulations, les contractions musculaires permettent de
rapprocher ces segments osseux. Le muscle triceps sural par exemple, formant le relief du
mollet, s'insère entre la partie inférieure du fémur et la partie postérieure du talon qu’on
appelle le calcanéum. Sa contraction va très logiquement entraîner une élévation du pied,
c'est-à-dire une extension du pied.

4*
Physiologie*de*l’exercice*–*B.*BARON*et*Pr.*P.*PELAYO*
4
Ce phénomène de contractions est macroscopiquement visible. On voit par exemple très
distinctement le triceps sural se raccourcir lorsqu’il entraîne une extension du pied sur la
jambe. C’est également le cas pour le biceps brachial lorsqu’il fléchit l'avant-bras sur le
bras.
Ces exemples correspondent aux contractions les plus classique durant lesquelles le
muscle se raccourci: c'est ce qu'on appelle une contraction concentrique.
Les autres modalités sont la contraction isométrique durant laquelle il n’y a pas de
déplacement segmentaire : on résiste à une charge sans qu’il n’y ait de mouvement. Et la
contraction excentrique : on ralenti une charge sans parvenir à l’immobiliser ou à la
soulever. Cette modalité est particulière car le muscle se contracte en s’étirant.
Si ce raccourcissement musculaire est macroscopiquement visible, c'est qu'il se passe
exactement la même chose microscopiquement au niveau de chaque fibre musculaire. Les
filaments fins et épais glissent les uns sur les autres sans qu’ils ne changent eux-mêmes de
longueur. Le sarcomère par contre passe d'une longueur d'environ 4 µm en position de
repos à une longueur de 2.7 µm lorsque la fibre musculaire contractée. Ce
raccourcissement microscopique de l’ensemble des sarcomères entraîne un
raccourcissement macroscopiquement visible permettant au muscle de générer une force
importante.
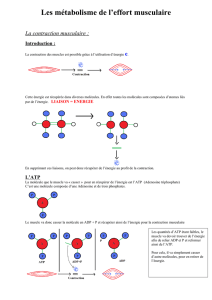
4. Approche biologique de la contraction musculaire.
Au niveau de la tête de myosine, on trouve une enzyme ayant la particularité d'hydrolyser
l'ATP. Cette enzyme est la myosine ATPase.
(Rappelons qu'une enzyme est une molécule permettant de faciliter une réaction chimique
et que l'hydrolyse de l'ATP permet de libérer de l'énergie comme nous le verrons dans le
chapitre réservé à la bioénergétique).
Lors de la contraction musculaire, la libération d'énergie induite par l'action de la myosine
ATPase permet le glissement entre les filaments d'actine et de myosine.

Physiologie*de*l’exercice*–*B.*BARON*et*Pr.*P.*PELAYO*
5*
5
Ce processus complexe se déroule en plusieurs étapes qui permettent de transformer une
énergie chimique en énergie mécanique à partir d'une décharge électrique.
La première étape correspond à l'arrivée du message nerveux sur la fibre musculaire. Son
point de départ se situe au niveau du cerveau, pour aller jusqu’à la fibre musculaire après
avoir transité le long de la moelle épinière. Le potentiel d'action émis par le système
nerveux arrive au niveau des tubules transverses de la cellule musculaire. C'est l’étape de
l'excitation musculaire.
(Les tubules transverses, également appelée tubules T, sont des invaginations du
sarcolemme délimitant le contenu de la cellule musculaire).
Le calcium (Ca2+) présent au niveau du réticulum sarcoplasmique va alors être libéré dans
le cytoplasme cellulaire. C’est l’étape du couplage excitation-contraction qui
correspond à l'ensemble des phénomènes permettant de transformer le signal nerveux en
un signal intracellulaire vers les fibres contractiles.
(Le réticulum sarcoplasmique est un compartiment intracellulaire qui stocke le calcium en
grande quantité).
Vient ensuite l’étape de la contraction musculaire proprement dite.
Le calcium libéré du réticulum sarcoplasmique se fixe sur le système troponine-
tropomyosine. Alors que celui-ci empêche habituellement l'interaction entre l’actine et la
myosine, l'arrivée du calcium lui fait perdre son pouvoir inhibiteur. L'actine et la myosine
se fixent alors. D'un point de vue chimique on peut représenter cette interaction par la
réaction :
actine + myosine ATPase → Actomyosine ATPase.
L'ATP est alors hydrolysé :
Actomyosine ATPase → Actomyosine + ADP + P + NRJ.
Le transfert d'énergie entraîne le mouvement entre les filaments fins et épais, par une
rotation de la tête de myosine. La fibre musculaire est alors contractée, le sarcomère est
raccourci.
Ce phénomène dure tant que la contraction de calcium est suffisante pour inhiber le
système troponine-tropomyosine. Lorsque la stimulation nerveuse s'arrête le calcium est
re-pompé vers le réticulum sarcoplasmique, le système troponine-tropomyosine reprend
son pouvoir inhibiteur. L'actine et la myosine restent alors séparées. C’est l’étape de la
décontraction. Il faudra attendre l'arrivée d'un nouveau potentiel d'action que la fibre
musculaire puisse à nouveau se contracter.
5. Différents types de cellules musculaires.
Lorsqu' on observe les fibres musculaires des sportifs spécialistes d'activités très intenses
et de courte durée (comme le sprint en athlétisme par exemple) après prélèvement par
biopsie musculaire, et qu'on les compare avec celles de spécialistes d’activités plus
longues (comme le marathon) des différences importantes apparaissent.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
1
/
33
100%