D1-ue5-Verkindt-Electroneuromyographie_Word

1
UE5 – Appareil locomoteur
C. VERKINDT
Date : 24/09/15 Plage horaire : 10h45-12h45
Promo : DCEM1 Enseignant : C. VERKINDT
Ronéistes :
BOUCHER Luc-Olivier
BOYER Aurélie
Bases physiologiques et techniques pour l’exploration
du SNP :
électro-neuro-myographie
I) Rappels
1. Rappels anatomiques
2. Genèse des potentiels d’action
3. Rappels anatomiques (suite)
II) Les déterminants de la détection des potentiels d’action
1. Buts de l’ENMG :
2. Dispositif d’enregistrement
3. Détection du signal
III) L’électroneuromyographie (ENMG)
1. EMG de surface
2. Les électrodes aiguilles
3. EMG de repos
4. Contraction volontaire maximale
5. Evaluation des unités motrices
6. Etude de la conduction motrice : neurographie motrice
7. Etude de la conduction sensitive : neurographie sensitive
8. Etude de la transmission neuromusculaire
9. Etude des réflexes

2
I. Rappels
1. Rappels anatomiques
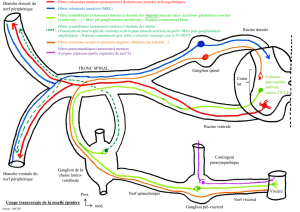
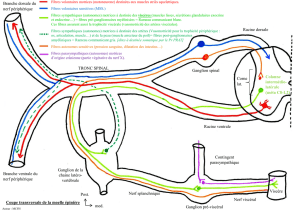
Des fibres afférentes (sensitives) provenant de la périphérie (muscles) rentrent dans la corne dorsale
de la moelle épinière. Ils présentent un corps cellulaire au niveau du ganglion spinal. Le neurone
bipolaire ramène l’information vers le SNC et les fibres efférentes (motoneurones de la ME) sont
des fibres motrices.
Les fibres efférentes sortent par les racines ventrales. Elles vont rejoindre la périphérie, en particulier
les motoneurones qui vont rejoindre les muscles squelettiques pour le SNS ou les muscles lisses pour
le SNA.
Les racines nerveuses qui sortent de la ME sont soit purement sensitives soit purement motrices, mais
au niveau des nerfs périphériques (ex : nerf spinal), elles sont mixtes, donc contiennent des fibres
motrices et sensitives. Très peu de nerfs sont purement moteurs ou sensitifs, on en trouve dans les
nerfs crâniens mais il n’y en a pas au niveau de la ME.
L’ENMG va permettre d’explorer tout ce qui se trouve en périphérie. On va explorer :
- Le bon fonctionnement de la transmission nerveuse sur les nerfs périphériques et / ou
- Le bon fonctionnement des effecteurs, en particulier les effecteurs musculaires avec la
transmission neuromusculaire et le fonctionnement du muscle.

3
2. Genèse de potentiels d’action
On a ici une portion d’axone, donc de fibre nerveuse. Un PA est une inversion de la polarité de la
membrane qui est due principalement à une entrée d’ions sodium. La membrane normalement
polarisée (potentiel de repos : charge + à l’intérieur et charge – à l’extérieure) à -70/-80 mV (pour un
neurone) va passer à +30mV.
Lorsque la dépolarisation atteint un certain seuil, elle provoque une inversion de polarité qui suit la
loi du tout ou rien. La propagation de ce PA se fait de proche en proche par l’ouverture des canaux
sodium voltage-dépendant. Elle va donc pouvoir progresser potentiellement des 2 côtés (si la
dépolarisation est appliquée au milieu d’un axone, la propagation du PA peut aller de chaque côté).
Ce n’est pas au niveau de la membrane axonale (donc pas au milieu de l’axone) que la conduction
physiologique se décide mais plutôt au niveau du corps cellulaire ou de l’extrémité dendritique pour
les neurones sensitifs. Donc, en conditions physiologiques, le PA ne se déplace que dans un sens. En
effet, il y a ouverture des canaux potassium, ce qui permet un rééquilibrage du potentiel avec une
sortie des ions K+ puis la repolarisation membranaire, avec parfois une hyperpolarisation transitoire
(période réfractaire).
En résumé, physiologiquement : PA → entrée ions sodium → +30mV → sortie d’ion K+ →
repolarisé.
Pour une fibre sensitive, la conduction orthodromique correspond au sens physiologique du
déplacement du PA, de la périphérie vers la ME (extrémité dendritique – corps cellulaire, puis corps
cellulaire – extrémité axonale). La conduction orthodromique dans les fibres motrices va de la ME
vers la périphérie (corps cellulaire – extrémité axonale). Une conduction qui va dans le sens inverse
du sens physiologique est une conduction antidromique.
3. Rappels anatomiques (2)

4
Une unité motrice est composée d’un motoneurone et de toutes les fibres musculaires que ce
motoneurone innerve. Ici on a un motoneurone de la corne ventrale de la ME. Chaque motoneurone
est à l’origine d’une unité motrice. Les unités motrices sont de tailles variables (5-6 fibres
musculaires dans les muscles oculaires, 1000-3000 dans les plus gros muscles de la cuisse par
exemple). Au niveau de chaque plaque motrice, on va avoir une dépolarisation du sarcolemme
provoquée par l’arrivée des PA qui va se propager sur toute la longueur de la fibre musculaire, d’où
contraction musculaire.
La gaine de myéline permet une conduction 20 fois plus rapide. De plus, plus le diamètre de la
fibre est important, plus la conduction est rapide.
II. Les déterminants de la détection des PA
On va utiliser des électrodes pour enregistrer l’activité électrique. On dispose les électrodes à
proximité de la zone que l’on veut explorer (fibres musculaire ou nerf). Ci-dessus, différents types
d’électrodes présentées (en rouge c’est le muscle et en vert la zone cutanée).

5
On positionne des électrodes de surface (sur la peau) avec une électrode active et une électrode de
référence, puis on capte un signal qui correspond à l’activité globale du muscle, on enregistre une
activité plus ou moins complexe. Plus on est distant du potentiel (du signal) plus on aura de difficulté
à capter ce dernier. Le PA (potentiel d’action) s’annule très vite avec la distance, donc nécessité
d’amplifier le signal. On peut également utiliser des électrodes aiguilles que l’on enfonce dans le
muscle.
Il y a des électrodes sous-cutanées qui permettent de se rapproche du site que l’on veut explorer, on
perdrait donc moins de signal. Ce sont des électrodes
in situ
qui ont l’avantage d’être très proches
du muscle que l’on veut explorer mais qui ont une surface d’enregistrement très faible.
On va jouer sur 3 facteurs :
- Distance de l’électrode par rapport à ce que l’on veut explorer. L’intensité du signal
recueilli diminue très rapidement avec la distance de l’électrode.
- Volume de recueil utile. Par exemple pour l’électrode aiguille cela correspond à un
petit cercle blanc sur le schéma ci-dessus. On ne capte que le signal au niveau du cercle
et on ignore ce qui se passe dans tout le reste du muscle. Ce volume de recueil utile est
donc déterminé par le type d’électrode utilisée. Donc volume de recueil utile augmente
→ signal plus global.
- Impédance du circuit électrique en kiloOhm (les résistances). On enlèvera tous les
poils, etc… qui pourraient gêner et/ou fausser le résultat.
Ces 3 paramètres vont permettre de déterminer la qualité du signal en ENMG.
III. L’Electro-neuro-myographie (ENMG)
L’ENMG va permettre l’exploration du SNP.
• Electromyographie : enregistrement de l’activité électrique musculaire qui peut être de
surface ou par électrode-aiguille.
• Etude de conduction des influx nerveux (vitesse de conduction, coupure de conduction…)
en positionnant les électrodes au dessus des nerfs.
• Techniques de stimulodétection (ex : explorer la voie réflexe) : exploration de la qualité
d’une partie du SNP ou d’une réponse musculaire avec une stimulation électrique.
Electrode de surface (sur la peau) → Distance élevée donc signal faible,
→ Volume recueil utile plus élevé donc signal plus global et plus
complexe.
Electrode aiguille (dans le muscle) → Distance faible donc signal fort,
→ Volume recueil utile plus petit donc signal plus ciblé et plus
restreint,
→ Augmentation de l’impédance car électrode fine donc
résistance importante.
1. Buts de l’ENMG
− Confirmer une hypothèse clinique : la technique ne fera jamais un diagnostic à notre
place, elle confirme un diagnostic. On ne voit que ce qu’on veut regarder, la machine ne dit
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
1
/
24
100%