Les oursins

Les oursins
I] Introduction
Les oursins font partie du phylum des échinodermes (7000 espèces toutes marines), et représente la classe des échinidés (900
espèces), qui sont deutérostomiens. La segmentation est dite holoblastique (totale), avec segmentation radiaire. Les
deutérostomiens ont la capacité de réguler des blastomères.
Ces échinides sont benthiques, et se retrouvent toujours à proximité des champs d'algues, du fait de leur régime
majoritairement herbivore. La nutrition se fait par le pôle oral, avec la lanterne d'Aristote (mastication) composée de 5 dents,
avec entre chaque dent une partie squelettique : le compas. Le système de locomotion est également en face orale, avec le
système de podias ambulacraire. Les piquants se retrouvent entre ces régions ambulacraires, on parle d'espace inter-
ambulacraire.
L'étude des oursins a permis de faire des découvertes importantes au niveau de la fécondation et du développement
embryonnaire. C'est un organisme modèle. Ils ont une fécondation et un développement externe, avec une grande production
de gamètes (107 pour la femelle et 1012 pour les mâles). Le développement est rapide et synchrone . Le génome a de plus été
totalement séquencé, permettant de tester l'intérêt de chaque gène pour le développement embryonnaire. Si la manipulation
génétique n'est pas aisée sur ces organismes, les manipulations telles que les transplantations et les dissociations cellulaires
(pendant les premières divisions) sont facilement réalisables. La production de gamète est saisonnière, et le temps de
génération trop long ne permet pas d'études génétiques (plusieurs années avant d'avoir un animal mature).
Morpholinos : oligonucléotides d'une 20aine de base, qui va se diriger contre un gène quelconque.
II] Développement embryonnaire
Les oursins sont gonochoriques, et expulsent leurs gamètes au niveau de la face aborale, par cinq orifices génitaux. On parle
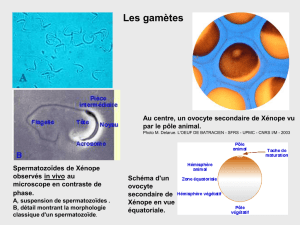
d'ovotides pour les gamètes femelles (orangés) et de spermatozoïdes pour les mâles (blanc).
1) Gamète mâle
Le spermatozoïdes est composé d'une tête contenant l'acrosome (contenant des hydrolases), d'un pool d'actine G), de la
chromatine dans le noyau (majeure partie de l'espace) et à la base du flagelles des mitochondries. Le flagelle est constitué de
microtubules et de dynéine (flagelle classique).
2) Gamète femelle
L'œuf est entourée de deux enveloppe, l'enveloppe vitelline collée à la membrane plasmique, et la gangue gélatineuse . On
retrouve des granules corticaux sous la membrane plasmique, avec un rôle dans la fécondation (activation de l'œuf).
La gangue gélatineuse aura un rôle dans le flottabilité de l'œuf, mais synthétisera aussi la resact (14 AA) qui attirera les
spermatozoïdes par chimiotactisme.
On retrouve une bande pigmentée (perpendiculaire à l'axe animal/végétatif), et au niveau du pôle animal, un canal traversera la
gangue, pour l'expulsion des globules polaires.
3) Fécondation
Quand les eaux se réchauffent, les gamètes sont expulsés en grande quantité dans l'eau. 48 heures après fécondation, on obtient
une larve pluteus, qui se déposera sur le fond au bout de 4 à 5 semaine (généralement en colonie).
Quand le spermatozoïde rencontre l'ovotide, on aura la réaction acrosomale : l'acrosome va hydrolyser la gangue par exocytose
des hydrolases, et le pool d'actine va polymériser pour percer la membrane vitelline avec la bindine contenu dans le filament
acrosomale. On aura un récepteur (spécifique pour chaque espèce) à la bindine dans la membrane vitelline. Les membrane
plasmiques fusionnent ensuite. Le contenu génétique se déversera alors dans l'ovotide, ainsi que toutes les structures du
spermatozoïde.
Dès la fécondation, le potentiel membranaire est rapidement inversé ( de -70 à 10 mV par entrée de cation par des canaux),
empêchant la polyspermie. Le potentiel initial sera rétabli en une dizaine de minutes. De plus, rapidement après la fécondation,

le calcium du réticulum est relargué dans l'ovotide, permettant l'activation de l'œuf par la réaction corticale : les granules
corticaux se vident entre la membrane plasmique et la membrane vitelline, créant une séparation entre membrane vitelline et
membrane plasmique (par des protéines coupant les liaisons entre ces membranes et création d'un gradient osmotique par des
sucres permettant l'entrée d'eau de mer) qu'on appelle la couche hyaline. La nouvelle structure soulevée s'appelle l'enveloppe
de fécondation, offrant une protection à l'œuf, et formant une barrière que ne peuvent franchir les spermatozoïdes (mise en
place après une dizaine de minutes).
On aura ensuite amphimixie : fusion des pronucléi au centre de l'œuf. Le noyau zygotique ainsi formé pourra commencer sont
développement par mitose rapides successives, avec activation du métabolisme.
4) Segmentation
La segmentation (ou clivage) holoblastique, radiaire, réductive commencera donc, la fin étant marqué par l'éclosion.
1. Clivage méridien → 2 cellules
2. Clivage méridien → 4 cellules
3. Clivage équatorial → 8 cellules
4. Clivage → 16 cellules non homogènes issues de mitoses asymétriques (4 micromères au pôle végétatif, au dessus 4
macromères, et 8 mésomères)
5. Clivage → 32 cellules (les 8 mésomères vont se diviser de façon équatoriale, générant 2 couches cellulaires An1 et
An2).
6. Clivage → 64 cellules avec différentes assises cellulaires perpendiculaire à l'axe animal/végétatif. On parle également
de stade morula.
7. Stade blastula (500 cellules) où chaque cellule développent un cil au pôle apical. Les cellules du pôle animal vont
avoir des cils plus longs, avec un rôle dans la direction.
La cavité centrale s'appelle blastocœle et on retrouve l'enveloppe de fécondation. La couche hyaline permet le maintient des
blastomères pendant le segmentation. Les cellules de la blastula font des jonctions serrées pour rester en contact.
On a pu, à l'aide de marquage, montré qu'au stade 64 cellules, les cellules veg 1 et veg 2 donneront le mésoderme, le reste des
veg 2 l'endoderme, et le reste des cellules l'ectoderme.
5) Gastrulation
Après formation de la plaque végétatif par aplatissement du pôle végétatif, des cellules vont exprimer de nouveaux gènes (et
donc protéines), et changer de forme. Elles perdent leur pôle basal, leur affinité avec la couche hyaline et les cellules
épithéliales et gagnent en affinité avec le blastocœle. Ces modification permet la sortie de cellules épithéliales (à l'aide de
filopodes) pour devenir cellules mésenchymateuses : on parle d'ingression.
Ces cellules mésenchymateuses vont former des spicules triradiés, pour servir de squelette de base à l'embryon et par
calcification former un tissu syncitial. Deux amas cellulaires oraux sont également formés, ainsi qu'une chaîne.
La gastrulation comment enfin par un léger iunfléchissement du pôle végétatif veg 2. Ces cellules s'invaginent dans le
blastocœle, et la poussée des cellules termine la gastrulation jusqu'à former l'archentéron. Le blastopore (première partie de
l'archentéron formée) donnera l'anus.
Lors de la première phase d'élongation de l'archentéron, il n'y a pas de division cellulaire, mais seulement un réarrangement
des cellules.
Lors de la seconde phase d'élongation, les cellule du bout de l'archentéron vont former un mésenchyme secondaire, qui va
émettre de longues extensions cytoplasmique : des filopodes. On parle de gastrula tardive. Les filopodes vont trouver un point
d'accroche, qui terminera la gastrulation en tractant l'archentéron vers la dépression stomodéale. Les cellules
mésenchymateuses secondaires se détachent et forment deux poches cœlomiques. Le sommet de l'archentéron fusionne avec
les cellules de l'ectoderme. Les cellules de l'ectoderme oral gardent une forme cuboïde, et les cellules de l'ectoderme aboral
vont s'allonger, formant un épiderme squameux).
6) Stade prisme
Des bandelettes ciliées vont permettre le déplacement, et l'attraction des particules nutritives vers le pôle oral. On aura des
ébauches d'organe. Cette larve s'allongera progressivement pour former la larve plutéus.

7) Larve plutéus
Les poches cœlomiques issues de la fin de la gastrulation se retrouvent autour de l’œsophage. Seul le cœlome gauche sera
impliqué dans le reste du développement : la poche cœlomique gauche se divise en 5 (pentaradiaire), formation de vestibule
(depuis ectoderme), pour former le rudiment (ou disque imaginal). A partir de ce rudiment, on aura naissance d'un oursin dit
juvénile (5-6 semaines).
Depuis le stade blastula à la fin du développement on passe d'une symétrie radiaire, à bilatérale, à dissymétrique droite/gauche,
et enfin symétrie pentaradiaire. On aura aussi formation graduelle du test calcaire dès la larve pluteus (l'alourdissement
permettant la fixation benthique).
III] Axe animal/végétatif
1) Existence de la polarité animale-végétative avant fécondation
On effectuer des coupures de l'œuf fécondé selon des plans différent. Une coupe transversale mènera à 2 larve quasi normales,
alors que la coupure équatoriale va donner d'une part une blastula permanente (issu du pôle animal) et une larve quasi normale
(issue du pôle végétatif). Ainsi, le pôle animal ne peut former que de l'ectoderme, alors que le pôle végétatif se suffit pour
former une larve normale.
Si on sépare les 4 cellules du stade 4, chaque cellule formera des larves. Au stade 8 cellules, une coupure méridionale induit
deux larves animales et une coupure équatoriale induit une larve normale, et une bloquée au stade blastula.
1
/
3
100%