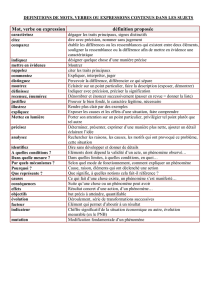
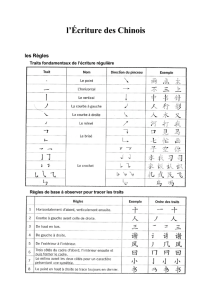
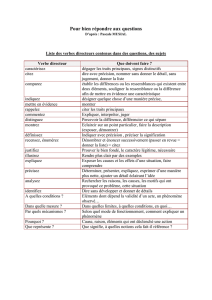
Thesis - Archive ouverte UNIGE
publicité

Thesis L'Europe du 3e millénaire avant notre ère et la question du Campaniforme : histoire des peuplements par l'étude des traits non métriques dentaires DESIDERI, Jocelyne Abstract La compréhension des mécanismes impliqués dans l'émergence du Campaniforme est abordée ici par l'étude des traits non métriques dentaires. Ce travail concerne cinq régions à occupation campaniforme (Espagne, France, Suisse, Bohême et Hongrie). Il s'agit de comprendre si la genèse du Campaniforme coïncide ou non avec un renouvellement de la population? Les cinq occupations analysées offrent un portrait riche et complexe de la société du 3e millénaire avant notre ère. L'implication du fonds local n'est pas le même selon les territoires étudiés. Deux sphères populationnelles distinctes ont été mises en évidence - l'une méridionale, l'autre orientale -, au sein desquelles les modalités de diffusion ne sont pas les mêmes. Dans la sphère méridionale, l'émergence du Campaniforme s'est produite par déplacements d'individus depuis la péninsule Ibérique. Dans la sphère orientale, une partie de la société locale se particularise en adoptant les traditions méridionales par un système d'exogamie. Reference DESIDERI, Jocelyne. L'Europe du 3e millénaire avant notre ère et la question du Campaniforme : histoire des peuplements par l'étude des traits non métriques dentaires. Thèse de doctorat : Univ. Genève, 2007, no. Sc. 3905 URN : urn:nbn:ch:unige-149257 DOI : 10.13097/archive-ouverte/unige:14925 Available at: http://archive-ouverte.unige.ch/unige:14925 Disclaimer: layout of this document may differ from the published version. UNIVERSITE DE GENEVE FACULTE DES SCIENCES Section de biologie Département d’anthropologie et d’écologie Laboratoire d’archéologie préhistorique et d’histoire des peuplements Professeure Marie Besse Professeur André Langaney L’Europe du 3e millénaire avant notre ère et la question du Campaniforme : histoire des peuplements par l’étude des traits non métriques dentaires THÈSE présentée à la Faculté des sciences de l’Université de Genève pour obtenir le grade de docteur ès sciences, mention archéologie préhistorique par Jocelyne DESIDERI de Grottammare (Italie) et Perly-Certoux (Genève) Thèse N° 3905 Genève 2007 Sommaire Remerciements ............................................................................................................................... 9 Morceaux choisis .......................................................................................................................... 17 Abstract ......................................................................................................................................... 19 Introduction ................................................................................................................................... 29 1. Le Campaniforme... un phénomène... une énigme ........................................................ 33 1.1. La situation précédant le Campaniforme ................................................................................. 33 1.2. Les faits ...................................................................................................................................... 34 1.2.1. Le matériel attribué au Campaniforme ................................................................................... 34 1.2.2. Les structures funéraires ........................................................................................................ 36 1.2.3. Les structures domestiques ................................................................................................... 39 1.2.4. La chronologie ....................................................................................................................... 41 1.3. Le Campaniforme hier ............................................................................................................... 42 1.4. Le Campaniforme aujourd’hui .................................................................................................. 44 1.5. Et quand l’individu associé à la culture Campaniforme s’exprime... .................................... 45 1.6. Un programme interdisciplinaire... un objectif commun ....................................................... 48 1.7. Les objectifs de cette recherche ............................................................................................... 49 1.8. L’essentiel en bref ..................................................................................................................... 51 2. Du groupe culturel à l’individu ........................................................................................... 53 2.1. Les régions sélectionnées ......................................................................................................... 53 2.2. Le domaine oriental : la Bohême .............................................................................................. 54 2.2.1. Le Campaniforme en Bohême : son insertion culturelle, spatiale et temporelle ........................ 54 2.2.2. Le Campaniforme en Bohême : les données de l’archéologie .................................................. 54 2.2.3. Le Campaniforme en Bohême : les données de l’anthropologie biologique .............................. 55 2.2.4. Le Campaniforme en Bohême : les échantillons étudiés .......................................................... 56 2.3. Le domaine oriental : la Hongrie .............................................................................................. 57 2.3.1. Le Campaniforme en Hongrie : son insertion culturelle, spatiale et temporelle ........................ 57 2.3.2. Le Campaniforme en Hongrie : les données de l’archéologie .................................................. 57 2.3.3. Le Campaniforme en Hongrie : les données de l’anthropologie biologique .............................. 58 2.3.4. Le Campaniforme en Hongrie : les échantillons étudiés .......................................................... 59 2.4. Le domaine méridional : le sud de la France ........................................................................... 60 2.4.1. Le Campaniforme dans le sud de la France : son insertion culturelle, spatiale et temporelle .... 60 2.4.2. Le Campaniforme dans le sud de la France : les données de l’archéologie .............................. 60 2.4.3. Le Campaniforme dans le sud de la France : les données de l’anthropologie biologique .......... 61 2.4.4. Le Campaniforme dans le sud de la France : les échantillons étudiés ...................................... 62 2.5. Le domaine méridional : le nord de l’Espagne ........................................................................ 63 2.5.1. Le Campaniforme dans le nord de l’Espagne : son insertion culturelle, spatiale et temporelle . 63 2.5.2. Le Campaniforme dans le nord de l’Espagne : les données de l’archéologie ............................ 63 2.5.3. Le Campaniforme dans le nord de l’Espagne : les données de l’anthropologie biologique ....... 64 2.5.4. Le Campaniforme dans le nord de l’Espagne : les échantillons étudiés .................................... 65 2.6. A l’articulation entre les deux domaines : la Suisse ............................................................... 66 2.6.1. Le Campaniforme en Suisse : son insertion culturelle, spatiale et temporelle .......................... 66 2.6.2. Le Campaniforme en Suisse : les données de l’archéologie ..................................................... 66 2.6.3. Le Campaniforme en Suisse : les données de l’anthropologie biologique ................................ 67 2.6.4. Le Campaniforme en Suisse : les échantillons étudiés ............................................................. 68 2.7. L’essentiel en bref ..................................................................................................................... 69 3. La terminologie dentaire ...................................................................................................... 71 3.1. L’anatomie de la dent ................................................................................................................ 71 3.2. Les dentitions ............................................................................................................................ 72 3.2.1. Les termes de positionnement ............................................................................................... 73 3.2.2. Les termes descriptifs morphologiques .................................................................................. 74 3.2.3. Les termes spécifiquement réservés aux dents antérieures ..................................................... 74 3.2.4. Les termes spécifiquement réservés aux dents postérieures ................................................... 75 3.3. Les systèmes de notation dentaire .......................................................................................... 76 3.4. Quelques définitions supplémentaires .................................................................................... 78 4. Les traits non métriques dentaires : des indicateurs des distances entre les populations ............................................................................................................................... 79 4.1. Un historique des recherches ................................................................................................... 79 4.2. Les traits non métriques dentaires .......................................................................................... 81 4.2.1. L’ontogénie dentaire .............................................................................................................. 81 4.2.1.1. Le développement dentaire .................................................................................................. 82 4.2.1.2. Les aspects moléculaires ..................................................................................................... 82 4.2.1.3. Les aspects biologiques ....................................................................................................... 85 4.2.2. Le mode de transmission des traits non métriques dentaires .................................................. 86 4.2.3. Les effets de l’environnement ................................................................................................ 88 4.2.4. L’héritabilité des traits non métriques dentaires ..................................................................... 90 4.3. La fiabilité du système d’enregistrement ................................................................................ 91 4.4. Le potentiel de ces variables dans les études entre populations .......................................... 93 4.5. L’essentiel en bref ..................................................................................................................... 96 5. De la dent au trait ................................................................................................................... 97 5.1. Les traits non métriques dentaires .......................................................................................... 97 5.1.1. La sélection des traits ............................................................................................................ 97 5.1.2. Les différents types de données dentaires ............................................................................ 101 5.2. L’acquisition des données ...................................................................................................... 105 5.2.1. La création d’une fiche d’enregistrement manuel ................................................................. 105 5.2.2. La création d’une base de données ...................................................................................... 105 5.3. L’évaluation de la concordance intra- et interobservateurs ................................................ 106 5.3.1. L’évaluation de la concordance dans la détermination de la dentition ................................... 106 5.3.2. L’évaluation de la concordance des modes d’observation des traits non métriques dentaires ............................................................................................................................. 109 5.3.2.1. L’évaluation de la discordance absolue ................................................................................. 110 5.3.2.2. L’évaluation de la concordance de l’observation des traits ....................................................... 110 5.3.2.3. L’évaluation de la discordance des graduations enregistrées .................................................... 111 5.3.2.4. Le pourcentage d’attribution de plus d’un stade de différence entre deux sessions ...................... 112 5.3.2.5. L’erreur moyenne brute ..................................................................................................... 113 5.3.2.6. Le degré de similarité des distributions ................................................................................ 114 5.3.2.7. La concordance de l’enregistrement des traits dentaires : un bilan ............................................ 115 5.4. L’essentiel en bref ................................................................................................................... 117 6. Le traitement préliminaire des données ....................................................................... 119 6.1. Les statistiques descriptives ................................................................................................... 119 6.1.1. Le traitement des traits non numériques .............................................................................. 119 6.1.2. Le décompte total des observations ..................................................................................... 120 6.1.3. La vérification de la transcription ......................................................................................... 121 6.1.4. La pertinence des variables observées .................................................................................. 121 6.1.4.1. L’état des variables observées pour la Bohême ...................................................................... 122 6.1.4.2. L’état des variables observées pour la Hongrie ...................................................................... 123 6.1.4.3. L’état des variables observées pour le sud de la France ........................................................... 124 6.1.4.4. L’état des variables observées pour le nord de l’Espagne ......................................................... 125 6.1.4.5. L’état des variables observées pour la Suisse ......................................................................... 126 6.1.4.5. L’état des variables observées : un bilan ............................................................................... 127 6.2. L’asymétrie des antimères ...................................................................................................... 128 6.2.1. Le degré de similarité des antimères .................................................................................... 129 6.2.1.1. Le résultat des corrélations entre les antimères pour la Bohême ............................................... 130 6.2.1.2. Le résultat des corrélations entre les antimères pour la Hongrie ................................................ 131 6.2.1.3. Le résultat des corrélations entre les antimères pour le sud de la France .................................... 132 6.2.1.4. Le résultat des corrélations entre les antimères pour le nord de l’Espagne .................................. 133 6.2.1.5. Le résultat des corrélations entre les antimères pour la Suisse .................................................. 134 6.2.1.6. Le résultat des corrélations entre les antimères : un bilan ........................................................ 135 6.2.2. Le traitement des antimères ................................................................................................. 136 6.2.2.1. Le traitement des antimères pour la Bohême ......................................................................... 138 6.2.2.2. Le traitement des antimères pour la Hongrie ......................................................................... 138 6.2.2.3. Le traitement des antimères pour le sud de la France .............................................................. 138 6.2.2.4. Le traitement des antimères pour le nord de l’Espagne ........................................................... 138 6.2.2.5. Le traitement des antimères pour la Suisse ........................................................................... 139 6.2.2.6. Le traitement des antimères : un bilan ................................................................................. 139 6.3. Les corrélations entre les traits .............................................................................................. 140 6.3.1. Les corrélations intradistrict ................................................................................................ 142 6.3.1.1. Les corrélations intradistrict pour la Bohême ......................................................................... 143 6.3.1.2. Les corrélations intradistrict pour la Hongrie ......................................................................... 143 6.3.1.3. Les corrélations intradistrict pour le sud de la France .............................................................. 143 6.3.1.4. Les corrélations intradistrict pour le nord de l’Espagne ........................................................... 143 6.3.1.5. Les corrélations intradistrict pour la Suisse ........................................................................... 144 6.3.1.6. Les corrélations intradistrict : un bilan ................................................................................. 144 6.3.2. Les corrélations interdistrict ................................................................................................ 145 6.3.2.1. Les corrélations interdistrict pour la Bohême ......................................................................... 146 6.3.2.2. Les corrélations interdistrict pour la Hongrie ......................................................................... 146 6.3.2.3. Les corrélations interdistrict pour le sud de la France .............................................................. 146 6.3.2.4. Les corrélations interdistrict pour le nord de l’Espagne ........................................................... 146 6.3.2.5. Les corrélations interdistrict pour la Suisse ........................................................................... 146 6.3.2.6. Les corrélations interdistrict : un bilan ................................................................................. 146 6.3.3. Les corrélations intertrait : tubercule de Carabelli (T24) - protostylide (T54) ......................... 147 6.3.3.1. Les corrélations intertrait pour la Bohême ............................................................................. 147 6.3.3.2. Les corrélations intertrait pour la Hongrie ............................................................................. 147 6.3.3.3. Les corrélations intertrait pour le sud de la France ................................................................. 148 6.3.3.4. Les corrélations intertrait pour le nord de l’Espagne ............................................................... 148 6.3.3.5. Les corrélations intertrait pour la Suisse ............................................................................... 148 6.3.3.6. Les corrélations intertrait : un bilan ..................................................................................... 148 6.4. Le dimorphisme sexuel ........................................................................................................... 149 6.4.1. Le test de la dépendance du sexe des individus dans l’expression des traits ......................... 150 6.4.1.1. Les résultats concernant le dimorphisme sexuel des traits pour la Bohême ................................. 151 6.4.1.2. Les résultats concernant le dimorphisme sexuel des traits pour la Suisse ................................... 151 6.4.1.3. Les résultats concernant le dimorphisme sexuel des traits : un bilan ......................................... 152 6.5. L’essentiel en bref ................................................................................................................... 153 7. L’analyse des données ......................................................................................................... 155 7.1. La procédure suivie ................................................................................................................. 155 7.1.1. Le calcul d’incidences populationnelles ................................................................................ 155 7.1.2. Le calcul de fréquences ajustées .......................................................................................... 155 7.1.3. La sélection des variables intégrées dans les analyses .......................................................... 158 7.1.4. Les analyses mobilisées ....................................................................................................... 159 7.2. L’analyse des ensembles ......................................................................................................... 160 7.2.1. Le domaine oriental ............................................................................................................. 160 7.2.1.1. La Bohême ...................................................................................................................... 160 7.2.1.1.1. La constitution des groupes testés ............................................................................... 160 7.2.1.1.2. Les incidences populationnelles .................................................................................. 161 7.2.1.1.3. Les variables mobilisées dans les analyses .................................................................... 162 7.2.1.1.4. Les analyses ............................................................................................................ 166 7.2.1.2. La Hongrie ...................................................................................................................... 167 7.2.1.2.1. La constitution des groupes testés ............................................................................... 167 7.2.1.2.2. Les incidences populationnelles .................................................................................. 167 7.2.1.2.3. Les variables mobilisées dans les analyses .................................................................... 168 7.2.1.2.4. Les analyses ............................................................................................................ 170 7.2.1.3. Les relations entre les ensembles du domaine oriental ............................................................ 171 7.2.1.3.1. La constitution des groupes testés ............................................................................... 171 7.2.1.3.2. Les variables mobilisées dans les analyses .................................................................... 172 7.2.1.3.3. Les analyses ............................................................................................................ 172 7.2.2. Le domaine méridional ........................................................................................................ 173 7.2.2.1. Le sud de la France ........................................................................................................... 173 7.2.2.1.1. La constitution des groupes testés ............................................................................... 173 7.2.2.1.2. Les incidences populationnelles .................................................................................. 173 7.2.2.1.3. Les variables mobilisées dans les analyses .................................................................... 174 7.2.2.1.4. Les analyses ............................................................................................................ 177 7.2.2.2. Le nord de l’Espagne ......................................................................................................... 178 7.2.2.2.1. La constitution des groupes testés ............................................................................... 178 7.2.2.2.2. Les incidences populationnelles .................................................................................. 178 7.2.2.2.3. Les variables mobilisées dans les analyses .................................................................... 179 7.2.2.2.4. Les analyses ............................................................................................................ 181 7.2.2.3. Les relations entre les ensembles du domaine méridional ........................................................ 182 7.2.2.3.1. La constitution des groupes testés ............................................................................... 182 7.2.2.3.2. Les variables mobilisées dans les analyses .................................................................... 183 7.2.2.3.3. Les analyses ............................................................................................................ 183 7.2.3. A l’articulation entre les deux domaines : la Suisse ............................................................... 184 7.2.3.1. La Suisse ........................................................................................................................ 184 7.2.3.1.1. La constitution des groupes testés ............................................................................... 184 7.2.3.1.2. Les incidences populationnelles .................................................................................. 184 7.2.3.1.3. Les variables mobilisées dans les analyses .................................................................... 185 7.2.3.1.4. Les analyses ............................................................................................................ 189 7.2.3.2. Les relations de la Suisse avec le domaine oriental ................................................................. 190 7.2.3.2.1. La constitution des groupes testés ............................................................................... 190 7.2.3.2.2. Les variables mobilisées dans les analyses .................................................................... 191 7.2.3.2.3. Les analyses ............................................................................................................ 192 7.2.3.3. Les relations de la Suisse avec le domaine méridional ............................................................. 193 7.2.3.3.1. La constitution des groupes testés ............................................................................... 193 7.2.3.3.2. Les variables mobilisées dans les analyses .................................................................... 194 7.2.3.3.3. Les analyses ............................................................................................................ 195 7.2.4. Les relations entre les populations campaniformes .............................................................. 196 7.2.4.1. La constitution des groupes testés ...................................................................................... 196 7.2.4.2. Les variables mobilisées dans les analyses ............................................................................ 197 7.2.4.3. Les analyses .................................................................................................................... 201 7.2.5. La Bohême : une analyse sexes séparés ................................................................................. 202 7.2.5.1. La constitution des groupes testés ...................................................................................... 202 7.2.5.2. Les incidences populationnelles .......................................................................................... 202 7.2.5.3. Les variables mobilisées dans les analyses ............................................................................ 203 7.2.5.4. Les analyses .................................................................................................................... 205 7.3. L’essentiel en bref ................................................................................................................... 207 8. De la discussion à l’interprétation .................................................................................. 211 8.1. Une discussion à l’échelle régionale ...................................................................................... 211 8.1.1. La Bohême ........................................................................................................................... 211 8.1.2. La Hongrie ........................................................................................................................... 213 8.1.3. Le sud de la France .............................................................................................................. 214 8.1.4. Le nord de L’Espagne ........................................................................................................... 215 8.1.5. La Suisse ............................................................................................................................. 216 8.2. Une discussion à grande échelle ............................................................................................ 217 8.3. Une proposition de dynamique des peuplements campaniformes ..................................... 219 Conclusions .................................................................................................................................. 223 De laboratoires en instituts : quelques souvenirs en images... ................................. 227 Bibliographie ............................................................................................................................... 229 Liste des figures ......................................................................................................................... 255 Liste des annexes ....................................................................................................................... 259 Liste des tables ........................................................................................................................... 261 Annexes et tables ................................................................................................................ CD-rom Remerciements Je tiens à remercier ici toutes les personnes qui ont largement contribué à rendre ma recherche des plus agréables. **** C’est avant tout à mes deux directeurs de thèse que je m’adresse pour m’avoir donné l’opportunité de réaliser ce travail, à la professeure Marie Besse, Département d’anthropologie et d’écologie, Université de Genève Chère Marie, je te suis tout d’abord infiniment reconnaissante de m’avoir confié ce sujet au sein du programme de recherche que tu conduis avec ton enthousiasme légendaire, ta rigueur exemplaire, ta bonne humeur d’une constance impressionnante et ton souci de bien-être de tes collaborateur/trices. Ensuite, je te remercie de m’avoir donné l’occasion d’évoluer à tes côtés, car j’ai pu bénéficier, non seulement d’un soutien de tous les instants, mais également d’une expérience enrichissante m’ayant permis de grandir scientifiquement. Enfin, tu m’as transmis ta passion pour le Campaniforme, qui plus d’une simple énigme, est un passionnant casse-tête de la préhistoire européenne ! au professeur André Langaney, Département d’anthropologie et d’écologie, Université de Genève Cher André, je te remercie d’avoir accepté de co-diriger cette recherche lorsque le département se trouvait sans professeur responsable en archéologie préhistorique car tu m’as donné la possibilité de l’accomplir. **** nsuite, c’est aux différents membres du jury qui ont l’amabilité de participer à l’évaluation de mon travail que va toute ma reconnaissance, à Anne-Marie Tillier, directrice de recherche au CNRS, Laboratoire d’Anthropologie des Populations du Passé, Université de Bordeaux, France au professeur Eric Crubézy, Laboratoire d’Anthropobiologie, Université Paul Sabatier, Toulouse, France au professeur Manuel Rojo Guerra, Département de préhistoire, Université de Valladolid, Espagne Chers professeurs, vos regards, vos remarques et vos critiques seront une aide précieuse pour la suite de mes recherches. **** Mes remerciements s’adressent ensuite aux différents directeurs du Département d’anthropologie et d’écologie de l’Université de Genève qui se sont succédés pendant ces cinq ans, m’ayant donné ainsi la possibilité de réaliser mon travail dans de bonnes conditions. 9 Je voudrais également remercier les personnes qui ont joué un rôle important dans l’orientation de mes recherches, notamment Christian Simon, qui a été à la tête du laboratoire de paléoanthropologie pendant près de vingt ans Ton décès prématuré, en 2000, a laissé un vide anthropologique certain. Christian, tu m’as donné l’opportunité de travailler sur la morphologie dentaire des populations campaniformes de Suisse occidentale. Tu me voyais déjà poursuivre cette étude en l’élargissant à l’échelle européenne... voilà qui est fait ! le professeur Alain Gallay, Département d’anthropologie et d’écologie, Université de Genève Cher Alain, je te serai éternellement reconnaissante pour ton enseignement, ton précieux soutien, tes conseils et ton enthousiasme. Ton jugement sur mes différents travaux a toujours été capital et je te remercie de l’avoir inlassablement réalisé avec un plaisir non dissimulé ! le professeur Christy G. Turner II, Département d’anthropologie, Arizona State University, Etats-Unis Vous avez eu l’amabilité de consacrer du temps à ma recherche et d’éveiller, par vos innombrables écrits, ma curiosité sur les tenants et aboutissants du monde fantastique de l’anthropologie dentaire. le professeur Alfredo Coppa, Département de biologie animale et humaine, Université de Roma La Sapienza, Italie Votre expérience en anthropologie dentaire a été fondamentale pour ma recherche. Vous avez toujours répondu présent à mes diverses interrogations. Mon travail a ainsi pu bénéficier de votre œil bienveillant ! **** Je ne saurais oublier d’adresser mes plus sincères remerciements aux différentes institutions qui ont contribué financièrement au bon déroulement de ma recherche, notamment le Fonds National de la Recherche Suisse pour la confiance placée à plusieurs reprises dans ce travail par l’attribution de diverses bourses (bourse de thèse dans le projet FNS 101412-100599 dirigé par Marie Besse et bourse de chercheur/se de la relève GE-112885) qui m’ont permis de m’y consacrer à temps complet. le Département d’anthropologie et d’écologie de l’Université de Genève pour avoir assumé la grande majorité de mes trajets à travers l’Europe, non seulement pour cette recherche, mais également dans le cadre de ma formation scientifique. la Société Suisse d’Anthropologie et l’Académie Suisse des Sciences Naturelles pour avoir financé plusieurs de mes voyages à travers l’Europe lors de la collecte des données. 10 Remerciements la Fondation Arditi pour l’intérêt dont vous avez fait preuve lors d’un travail préliminaire, mon mémoire de master, m’encourageant ainsi à poursuivre dans cette voie (prix Arditi 2002). **** J’ai eu le privilège d’évoluer au sein de deux équipes pendant ma recherche. C’est à l’ensemble de ces personnes que je souhaiterais dire merci - lesquelles, plus que de simples collègues, sont des vrai(e)s ami(e)s -, en particulier à l’équipe du 3e millénaire - nommée dans l’intimité les ami/es de la cloche -, notamment à Marie Besse, Louise-Sylvie Bourlon, Florence Cattin, Martine Piguet, et enfin Robin Furestier, venu viriliser une équipe - initialement - exclusivement féminine ! Ce fut un sincère plaisir et une réelle stimulation d’être intégrée dans une équipe interdisciplinaire telle que celle-ci et de travailler à vos côtés pendant ces cinq belles années. à mes collègues anthropologues d’hier et d’aujourd’hui, mais plus précisément à Christiane Kramar et Geneviève Perréard Lopreno - collègues et amies de toujours au sein de la famille des paléo... quoi ? ... anthropologues, ou bien ! Le bureau de l’amitié - nommé également l’annexe du jardin botanique - a été un lieu privilégié d’échanges, de formation et de réflexion d’une valeur unique, merci les filles ! **** Avoir choisi un sujet européen m’a permis de visiter un certain nombre d’instituts, de départements et de musées. Et c’est à l’ensemble des personnes rencontrées lors de mes différents séjours que je souhaite exprimer toute ma gratitude. Vous m’avez ouvert les portes de vos institutions - et parfois à plusieurs reprises - en me donnant accès à vos splendides collections, vous m’avez intégrée dans vos laboratoires, vous m’avez transmis des documents inédits. Merci pour ces rencontres et ces expériences enrichissantes tant au niveau humain que professionnel ! Ainsi, par ordre alphabétique, les diverses institutions qui m’ont accueillie : • Anthropologisches Forschungsinstitut, Aesch, Suisse • Département d’anthropologie, National Museum, Prague, République tchèque • Arcadia, Instituto de Promocion Cultural, Université de Valladolid, Espagne • Département d’anthropologie, Université de Szeged, Hongrie • • Aquincum Museum, Budapest, Hongrie • Département d’anthropologie, Musée d’histoire naturelle hongrois, Budapest, Hongrie Département d’histoire et de philosophie de l’Université de Alcalá de Henares, Espagne • Département de préhistoire, Université de Valladolid, Espagne 11 • Laboratoire d’Anthropologie des Populations du Passé, Université de Bordeaux, France • Laboratoire d’Anthropologie, Faculté de Médecine secteur Nord, Université de la Méditerranée, Marseille, France Museo Arqueológico de Burgos, Espagne • • Museo Arqueológico de Valladolid, Espagne • Museo Arqueológico Provincial d’Avila, Espagne • Museo Arqueológico Provincial de Palencia, Espagne • Service d'Anthropologie Biologique, Faculté de Médecine, Université de la Méditerranée, Marseille, France Et, toujours par ordre alphabétique, toutes les personnes que j’ai eu l’honneur de rencontrer et sans lesquelles cette recherche n’aurait pu être ce qu’elle est aujourd’hui. Merci à vous tou(te)s, notamment à Yann Ardagna à Aude Civetta Service d'Anthropologie Biologique, Faculté de Médecine, Université de la Méditerranée, Marseille, France Service d'Anthropologie Biologique, Faculté de Médecine, Université de la Méditerranée, Marseille, France à Isabelle Aymard à Patrice Courtaud Service d'Anthropologie Biologique, Faculté de Médecine, Université de la Méditerranée, Marseille, France Laboratoire d’Anthropologie des Populations du Passé, Université de Bordeaux, France à la professeure Rosa Barroso Département d’histoire et de philosophie de l’Université de Alcalá de Henares, Espagne à Gilles Boetsch directeur de recherche au CNRS, Service d'Anthropologie Biologique, Faculté de Médecine, Université de la Méditerranée, Marseille, France à Claude Bouville Service d'Anthropologie Biologique, Faculté de Médecine, Université de la Méditerranée, Marseille, France à la professeure Primitiva Bueno Ramirez Département d’histoire et de philosophie de l’Université de Alcalá de Henares, Espagne à Belén Castillo directrice du Museo Arqueológico de Burgos, Espagne à Viktor Cerny Institut d’archéologie, Academy of Sciences, Prague, République tchèque 12 à Coralie Demangeot Laboratoire d’Anthropologie des Populations du Passé, Université de Bordeaux, France à William Devriendt Service d'Anthropologie Biologique, Faculté de Médecine, Université de la Méditerranée, Marseille, France à la professeure Miluse Dobisikova directrice du Département d’anthropologie, National Museum, Prague, République tchèque à Eric Drenth Rijksdienst Voor het Oudheikundig Bouemonderzoek, Amersfoort, Hollande à Henri Duday directeur de recherche au CNRS, Laboratoire d’Anthropologie des Populations du Passé, Université de Bordeaux, France à Petr Dvorak Městské Muzeum, Brno, République tchèque à Anna Endrödi directrice de l’Aquincum Museum, Budapest, Hongrie Remerciements à José Francisco Fabian Garcia à Laure Salanova Institut d’archéologie, Avila, Espagne CNRS - Archéologies et Sciences de l'Antiquité, Maison René Ginouvès d'Archéologie et d'Ethnologie, Nanterre, France à Inigo Garcia-Martinez de Lagran Arcadia, Instituto de Promocion Université de Valladolid, Espagne Cultural, à Rafael Garrido-Pena Arcadia, Instituto de Promocion Université de Valladolid, Espagne Cultural, à Michel Signoli Service d'Anthropologie Biologique, Faculté de Médecine, Université de la Méditerranée, Marseille, France au professeur Eugen Strouhal à Gerhardt Hotz Institut d’histoire de la médecine et de langues étrangères, Université de Saint-Charles, Prague, République tchèque Naturhistorisches Museum, Bâle, Suisse au professeur Pedro Jiménez Sanz Département d’histoire et de philosophie de l’Université de Alcalá de Henares, Espagne à Yaramila Tcheremissinoff au professeur Bruno Kaufmann INRAP Toulouse et Centre d’Anthropologie, Toulouse, France Anthropologisches Forschungsinstitut, Aesch à Anne-Marie Tillier à Olivier Lemercier directrice de recherche au CNRS, Laboratoire d’Anthropologie des Populations du Passé, Université de Bordeaux, France Département d'Histoire de l'Art et Archéologie, Université de Bourgogne, Dijon, France à Jan Turek à Eric Mahieu Département d’archéologie, Université de SaintCharles, Prague, République tchèque Actilia multimédia, Theix, France à la professeure Antonia Marcsik à Magdalena Turkova Département d’anthropologie, Université de Szeged, Hongrie à Maria Mariné Isidro à Tanya Ulding directrice du Museo Provincial d’Avila, Espagne à Sergio Moral Osteo-Archäologie-Service, Aesch, Suisse à Zita Van der Beek Institut d’archéologie, Burgos, Espagne Oxford, Grande Bretagne à la professeure Ildiko Pap à Antonio Vázquez Cuesta Département d’anthropologie, Musée d’histoire naturelle hongrois, Budapest, Hongrie à Christina Papageorgopoulou Département d’histoire et de philosophie de l’Université de Alcalá de Henares, Espagne à Petr Veleminsky Anthropologisches Forschungsinstitut, Aesch, Suisse à Javier Pérez Département d’anthropologie, Museum, Prague, République tchèque National à Joël Vital directeur du Museo Arqueológico Provincial de Palencia, Espagne à Laszlo Remeneyi CNRS - Centre d'Archéologie Préhistorique, Valence, France à Piotr Wlodarczak Aquincum Museum, Budapest, Hongrie Institut archéologie et d’ethnologie, Cracovie, Pologne au professeur Manuel Rojo Guerra Département de préhistoire, Valladolid, Espagne Institut d’archéologie, Academy of Science, Prague, République tchèque Université de 13 mais également à tous les membres de l’association Archéologie et gobelets et à nos rencontres annuelles enrichissantes à travers l’Europe. **** Je souhaite aussi remercier un certain nombre de personnes, qui, généralement, reste dans l’ombre, mais dont l’aide et le soutien ont été indispensables pour la réalisation de ma recherche. Un merci particulier aux différents membres du PAT du département d’anthropologie et d’écologie, notamment au chef suprême de la Cafête Jean Gabriel Elia, aux informaticiens Georges Puissant, David Roessli et Stéphane Weber, aux techniciens Jacques Koerber et Matteo Gios, aux secrétaires Leila Gaudé et Marisa Andosilla, aux bibliothécaires Marie-Noëlle Lahouze Davaud et Valérie Mirault, aux photographes Micheline Vautravers et Elvyre Martinez pour votre grande patience… toujours prêts à résoudre les nombreux problèmes existentiels - et de la plus haute importance - auxquels j’ai été confrontée. à mes collègues du département d’anthropologie et d’écologie, notamment à Jean-François Buard, Elena Burri, Mathias Currat, Suzanne Eades, Viktoria Fisher, Loic JammetReynal, Sylvain Ozainne, Estella Sim Poloni, Caroline Robion-Brunner, Giulia Willig pour votre précieuse aide, vos judicieux conseils et pour avoir répondu, à un moment ou à un autre, à mes différentes interrogations. à Cécile Delhumeau, docteur en génie biologique et médical, Centre Médical Universitaire, Université de Genève à Magali Schweizer, docteur en paléontologie, Geologisches Institut, Université de Zurich pour votre savoir éclairé concernant la manipulation des données qualitatives. à mes revieweuses privilégiées, notamment à Marie Besse, Jasmine Desideri Oberson et Christiane Kramar pour avoir accepté cette lourde et ingrate tâche, pour la pertinence de vos critiques et pour votre sens de la rhétorique. Toutes trois ont permis de rendre ce travail bien plus agréable à lire ! **** 14 Remerciements mais aussi à tou(te)s mes collègues ayant hanté ou qui hantent toujours les couloirs du département d’anthropologie et d’écologie pour avoir partagé ces instants inestimables qu’est la vie au dep ! à mes petits camarades de la Comédie, avec lesquels j’ai passé d’innombrables soirées à refaire le monde et qui m’ont permis de relativiser, en particulier, à Aurélie, qui m’a donné un sacré coup de main dans l’informatisation de mes données. à mes amis du Popocatépetl pour les mille et un repas, discussions hautement philosophiques et bonne humeur que l’on a partagé des années durant... à mes amis pour avoir supporté et encouragé l’éternelle étudiante, ça y est c’est fait, l’éternelle étudiante n’est plus ! enfin et non des moindres... ... au clan Desideri et agréés pour leur soutien sans limites… GRAZIE ! **** ...mille pardons à tous ceux qui auraient dû apparaître ici et qui ont été oubliés... par écrit... uniquement, je vous l’assure !! 15 Morceaux choisis Période de transition entre la fin des temps néolithiques et les débuts de l’Age du bronze, le « Chalcolithique » constitue, dans l’évolution des sociétés, une période à la fois capitale et ambiguë. Capitale parce qu’elle est l’étape au cours de laquelle apparaissent de nouvelles avancées techniques, une production intensifiée, des échanges organisés sur de larges espaces, un accroissement des différences sociales, un cadre architectural où l’ostentation vient s’afficher aux côtés de l’ordinaire, le poids grandissant des modes idéologiques. Ambiguë parce qu’une fois ces concepts énoncés, il n’est pas aisé de décortiquer, au sein de chaque culture chalcolithique, les mécanismes de leur imbrication, le rôle de chacun d’eux dans l’amorce du processus de « complexité », une définition claire du fonctionnement social et spirituel des cultures concernées. On aura d’emblée compris que le renouvellement qui caractérise cette période joue autant au niveau de la production économique qu’à celui des relations sociales et du fonctionnement idéologique. Apprécier la part de chacun de ces facteurs dans une perspective systémique n’est pas chose aisée. (J. Guilaine 2007, p. 5) La question campaniforme continuera longtemps à figurer parmi les plus irritantes de la préhistoire européenne. (C. Jeunesse 1998, p. 418) ... malgré tant d’incertitudes, [elle] est l’une des plus grandes de l’histoire européenne. (R. Riquet, J. Guilaine et A. Coffyn 1963, p. 121) Les limites que rencontrent nos tentatives de restitution du passé ne doivent pas faire oublier les succès incontestables de la recherche. Les travaux des préhistoriens débouchent, malgré leurs imperfections, sur des acquis nourrissants, un savoir de plus en plus riche ; ils prouvent ainsi que l’histoire en général, et la préhistoire en particulier, font pleinement partie du domaine scientifique. Et surtout elles ne doivent pas nous priver du plaisir de rêver à nos plus lointains parents. (A. Gallay 2006, p. 13) 17 Abstract The Bell Beaker complex is defined, above all, by a ceramic style widespread across Europe during the 3rd millennium BC. In contrast to the preceding period, it represents an entity distinguished from other archaeological manifestations by its material culture, funeral practices and mode of diffusion. Its particularly vast geographic distribution has provoked different interpretations : a unique population invading Europe, the long-distance exchange of prestige goods, and the absence of a Bell Beaker population with only the diffusion of its cultural components. Osteoarchaeology makes it possible to test the first of these hypotheses, which proposes the diffusion of a culture by population displacement. The pertinent elements on the human skeleton must thus be identified for analysis. Here, the choice was made to analyze non-metric dental traits, anatomic variations observed on permanent and deciduous dentition. These are, in general, traits that are present or absent, or which reflect different phases of development. They can be positive structures, such as the presence of tubercles or ridges, or negative, such as grooves. Such traits can also vary in the number, position and size of cusps and roots. Based on results obtained from studies of archaeological material, the regions selected to test this hypothesis are the Czech Republic, Hungary, Switzerland, France and Spain. Chapter 1 The first chapter presents the basis of this research : the Bell Beaker complex. We are interested in previous interpretations and current thought with respect to the definition of the Bell Beaker complex. The present research is also placed within an interdisciplinary program addressing the population history of the 3rd mill. BC. Finally, • The Bell Beaker complex was established on a Europe-wide scale, with quite different preceding local substrates. Between the uniform Eastern pre-Bell Beaker and the Western heterogeneity, data are difficult to compare. • The Bell Beaker complex is characterized by material including decorated pottery - the common denominator of this culture -, common ware ceramics varying by region, diverse artefacts including archers’ armbands, tanged daggers, Palmela points and buttons with V perforation, and quite beautiful anthropomorphic steles. • While these artefacts unite the Bell Beaker complex, the incontestable diversity of its funerary and domestic structures blurs the overall uniformity. From the individual grave, re-use, collective tombs and incineration, the funeral practices of the Bell Beaker reveal its complexity. Similarly, variability in domestic structures, whether with respect to construction - on posts or dry-stone -, form - circular, oval, rectangular -, or location - near or far from their substrate -, is clear. • The Bell Beaker complex developed during the 3rd mill. BC, between 2900 and 1800 BC, and it is possible to demonstrate a southwest-northeast gradient for its emergence. • Initial explanations concerning the emergence of the Bell Beaker were primarily based on the panEuropean character of certain types of artefacts and vary between population movement, exchange of goods and exchange of ideas. • Today, research focuses on divergences, analyzing vast territories using specific instruments. The least exceptional elements of the material culture, such as common ware ceramics and lithics, express and demonstrate a clear dichotomy in the Bell Beaker complex that opposes the eastern and western regions of this phenomenon. 19 • Anthropological studies are not lacking, from the specialized study to a pan-European scale, from analysis of cranial to the chemical composition of bones. Mobility, - partial, total, or varying depending on the approached employed, is often associated with the emergence of the Bell Beaker complex. The specific cranial morphology of Bell Beaker individuals forms the basis for this : a brachycephalic skull with flattened occipital. Such morphology has long been considered as a definitive proof for population movements at the beginning of the Bell Beaker period. Today, other, less subjective, approaches, such as isotopic geochemistry of bone and non-metric dental traits, are being applied. • This research has been integrated within an interdisciplinary program aimed at understanding the mechanisms for the emergence of the Bell Beaker phenomenon by using four complementary axes : the typology and chronology of the common ware ceramic, territorial occupation, metallurgy and dental anthropology. These different elements define the main objective of this research : was there a rupture or continuity in populations during the appearance of the Bell Beaker complex? To respond to this question, five regions Bohemia, Hungary, southern France, northern Spain and Switzerland - were selected, representing the majority of the variability present in the Bell Beaker, totalling more than 2’000 individuals, 530 traits and 255’000 observations. Chapter 2 This chapter presents both the different regions and samples integrated in this research. While far from representing the entire range of variability in the Bell Beaker complex, an attempt has been made to cover a large portion of this variability. Five regions representing five different portraits of the Bell Beaker occupation during the 3rd mill. BC have been chosen. • The eastern domain : Bohemia This region, often considered as one of the major actors in Bell Beaker population history, represents the eastern-type occupation. The Bell Beaker developed during the second half of the 3rd mill. BC, during the final phase of the Eneolithic, in an intermediate position between the Corded Ware and the Unetice cultures. Archaeological data demonstrate the great affinities of the Bell Beaker with the two cultures preceding and succeeding it, as much with respect to funerary practices as artefacts themselves. Analysis of human remains, in contrast, suggests a certain degree of mobility of individuals that differs depending on the approach employed. A sample of more than 500 individuals and 64 sites from the northern and central regions of Bohemia has been selected. They belong to the Corded Ware, Bell Beaker and Unetice (Early Bronze Age) cultures. • The eastern domain : Hungary The Hungarian Bell Beaker occupation - the Csepel-Bell-Beaker - is the only one in the eastern domain outside the zone of the Corded Ware substrate. It is integrated within the middle phase of the Early Bronze Age (BA II), during the second half of the 3rd mill. BC, and is limited to the region around Budapest. Different interpretations have been advanced to explain the limited intrusion of the Bell Beaker complex in Hungary. For some, the origin of this complex would have resulted from the migration of small groups from the Czech Republic; for others, it would have been due to an internal development of Early Bronze Age society. The rare anthropological data plead in favour of a population renewal without excluding the possibility of contacts with contemporaneous populations. Here, a corpus has been composed of around a hundred individuals and 15 sites located in the northcentral and southern regions of Hungary. These assemblages are attributed to the Csepel-Bell Beaker and to contemporaneous (Obedà-Pitvaros, Nyirseg and Somogyvar-Vinkovci) or succeeding (Nagyrev, Perjamos) cultures. 20 Abstract • The southern domain : Southern France The end of the Neolithic, marked by the emergence of a multitude of distinct and geographically limited cultural groups, is divided into two phases : the Final Neolithic and the Chalcolithic. The Bell Beaker was established in irregular manner across the region considered during the latter phase and, in some cases, coexisted with local cultures. It developed during the first half of the 3rd mill. BC. Archaeological data generally reveal two phases of Bell Beaker occupation. The first marks a clear rupture with local traditions, the second a period of integration and cohabitation. The complicated funerary contexts in this region have probably not led to many anthropological studies. The sparse data do not permit interpretation in terms of population history. The sample consists of around 700 individuals and five sites mostly located in the eastern part of the region. The assemblages are chronologically situated between the Final Neolithic/Chalcolithic and the Early Bronze Age. • The southern domain : Northern Spain The Iberian Peninsula is a significant region for understanding the Bell Beaker phenomenon. The Meseta is a territory rich in discoveries. The Bell Beaker developed during the second half of the 3rd mill. BC, during the recent phase of the Chalcolithic and coexisted with it until the emergence of the Early Bronze Age. Archaeological data suggest that the emergence of the Bell Beaker was associated with societal transformations at the end of the Neolithic by the exchange of socially valuable goods. From an anthropological point of view, no element suggests a renewal or displacement of populations at the origins of the Bell Beaker; however, studies have not specifically addressed this issue and concern primarily the effects of environment on individuals. The sample here consists of around 180 individuals and 14 sites located in the northern part of the Meseta. The assemblages are attributed to the Final Neolithic, Chalcolithic, Bell Beaker and Bronze Age. • At the articulation between the two domains : Switzerland The Swiss territory is located in an intermediary position between the southern and eastern domains. Archaeological material, unequally distributed across Switzerland, demonstrates an east-west duality of traditions in different cultural areas. The Bell Beaker appeared during the second half of the 3rd mill. BC, at the end of the Neolithic, and was succeeded by the emergence of the Early Bronze Age. Based on the archaeological data, the origin of the Bell Beaker in Switzerland could have been the result of more or less marked influences from both the southern domain (ideological influences) and the eastern domain (population migration). Different anthropological studies note a high degree of homogeneity in pre-Bell Beaker populations. By contrast, results for successive populations are more uncertain, sometimes suggesting population continuity, sometimes invoking population renewal. The sample consists of more than 520 individuals and eight sites mainly located in the western part of Switzerland. This region presents a sequence without a major hiatus from the 5th to 3rd mill. BC, and samples were selected that are chronologically situated between the Middle Neolithic and the Early Bronze Age. As can be seen, the data are not comparable from one region to the other. The multiple facets of the Bell Beaker phenomenon prevent a single interpretation for its origin; anthropological data illustrate rather the complexification of the Bell Beaker phenomenon. Chapter 3 The objective of this chapter is to present a lexicon of terms specific to this research. It was intentionally decided to present these terms in the form of text, rather than a glossary, to facilitate reading. 21 Chapter 4 At the conclusion of this chapter, we saw that dental traits are a valuable tool for understanding relationships between populations. On the one hand, dentition is one of the most resistant elements of the skeleton, and thus often the best preserved. On the other hand, non-metric dental traits can be observed on living people, and research on genetic determinism is facilitated by direct analysis of similar subjects or twins. The reference to modern populations finds its significance here. Scientific progress has also contributed to improving understanding of the mechanisms and processes involved in odontogeny. • First, research has established that dental development, and in particular that of dental morphology, is highly subject to genetic control. Several studies have demonstrated that the development of the dentition is controlled by a certain number of genes that act in different places and intervene at distinct moments of its formation. While no trait follows a simple mode of inheritance, they are not as complex as those of models of polygenic inheritance in which many genes are involved. In addition, although we cannot deny a probable influence of environmental factors, we also cannot contest the fact that their effects are minor. Finally, studies based on genetic determinism have demonstrated that most of these traits seem to possess a large hereditary component. • Next, we have seen that the viability of the system of observation is an essential element in the validation of data. In effect, the lack of standardization of data can sometimes lead to entirely different results. The subjectivity of the observation has often been a determining element in the criticism of the study of dental traits. But at present, it is possible to move past this obstacle, by using dental casts and precise definitions of the different variables, and by eliminating traits that could pose possible problems of standardization by testing not only their own accordance, but also that between different observers. • Finally, studies on the value of non-metric dental traits when applied to modern populations are clearly reassuring. Comparison of results of dental morphology with different estimators, such as geographic proximity, linguistics and genetic data, have reinforced the idea that these variables are good indicators of biological distance between populations. Today, research on non-metric dental traits continues and leads to a better understanding of these variables. Moreover, we can only rejoice in the accordance of results between dental anthropology and genetics since it seems that dental data on northern Asia, southeast Asia and the Americas are generally in excellent agreement with those from single genes (Cavalli-Sforza et al. 1994, p. 317). Even if some aspects should be better mastered, notably the genes responsible for the development of these variables, to cite only a single example, the genetic determinism underlying the expression of dental traits allows them to be used for comparative studies between populations. Chapter 5 This chapter discusses the selection of variables used for this research, their mode of acquisition and evaluation of the accordance of the data. • During the first phase, 111 dichotomous or graduated traits were selected, representing 530 possible observations. For permanent dentition, we have retained the traits proposed by the ASU system (Turner et al. 1991) and the FU-DTS system (Alt 1997). For deciduous dentition, we have selected the traits defined by K. Hanihara (1963), P.W. Sciulli (1977) and K.D. Jorgensen (1956). In addition, some of the traits recorded on permanent molars could be observed on second deciduous molars. 22 Abstract In most cases, we have retained the expressions proposed by the different authors and recording systems. 11 traits were, however, modified because the graduations presented were deemed difficult to identify. • During the second phase, we focused on the mode of data acquisition. Manual data recording forms were used as well as a data management system to permit the easy manipulations and rapid transfer to data for use in different computer programs. • Finally, we measured the accordance between the determination of teeth and the observations of non-metric dental traits. Evaluation of the accordance of determination of the dentition was carried out even if the principal objective of the method was the correct cultural attribution of a sample essentially comprised of isolated teeth (dolmen of Peirières, Villedubert, Aude, France). We were able to show evidence for different kinds of errors (notation, lateralization, interarcade, interdistrict and interdentition). Disagreements were not numerous. The most common error was that of incorrect identification of the number attributed during excavation (n=79); teeth with this type of error were eliminated. Evaluation of the accordance of modes of observation of non-metric dental traits yielded interesting results. Intra-observer results indicate that differences in gradations between recording sessions are minimal and thus negligible. We nevertheless eliminated six variables observed on deciduous dentitions that were deemed difficult to observe. By contrast, differences between observers were more significant and confirm the difficulty of using data that have not been directly collected by the researcher. Chapter 6 This chapter presents the preliminary treatment of dental observations. This treatment entailed various aspects, such as verification, refinement and optimization of data prior to analysis. It forms a phase of delicate data manipulation, involving important decisions. These are summarized as follows : • Descriptive statistics This part concerns the descriptive phase of raw data. Initially, the variables were identified alphabetically in order to treat them statistically (T11 to T53 on permanent dentition). Next, an inventory of observations by dentition was made for each region. More than 255’000 observations were recorded on more than 2000 individuals. The next step was to verify the transcription of observations into the computer database (typing errors, aberrant gradations, discrepancies in recording, etc.) in order to correct such errors. Finally, the relevance of the variables observed was evaluated. Traits with an insufficient sample size were eliminated. This suppression concerned only variables observed on deciduous dentition for Bohemia, Hungary and Northern Spain. Data which was constant was also eliminated during this step. • Asymmetry of antimeres This step was essential since it involved the transformation of bilateral observations in unique data in order to eliminate redundant information provided by the antimeres. This first consisted in measuring the degree of similarity of expression of dental traits observed on controlateral teeth, and this for each region. We note that the relationships and intensity of antimeric expressions depend on the specificity of the samples studied. Thus, best results were obtained for Switzerland and Bohemia, comprising in majority individual subjects. Since non-correlated traits varied from one region to another, we have chosen to consider interdependent the antimeric expressions of the assemblage of variables. Next, treatment of bilateral data was carried out. Among the different methods used, two were retained : lateral count which retains only one antimere (the left arbitrarily selected here) and individual count which retains only the highest expression and preference. The selection of one or the other of these methods took place separately : 1) As a function of the type of variable : six traits - which are most often a form or a position - were subjected to a specific treatment. The individual count was used for T9, T11 (after dichotomization) and 23 T55-56 (after fusion of these two traits and a new gradation), observed on permanent dentition. Lateral count was used for T53 on permanent dentition, T3, T11 and T36 on deciduous dentition. These decisions were applied for all populations studied. 2) As a function of the state of the samples : Bohemia and Hungary were subjected to treatment by individual count. The other three regions, presenting composite situations, were treated according to the specificity of the sites : individual count for sites permitting individualization of the subjects and lateral count for those with mostly isolated teeth. • Correlation between traits This fraction concerned evaluation of relationships between variables for all populations. We focused on three types of associations : 1) Intradistrict correlations : Variables recorded were verified that they followed the expected model termed gradient of expression or morphogenetic field (if a trait is present on one member of the dental class, it is highly probable that it will be observed on all of the group with an increase in mesio-distal expression). Results demonstrate that traits not following this model vary between the populations studied. For reasons of sample representativeness, we chose only a trait not conforming to the model if the results of two major regions (CZ and CH) or a major region and at least two minor regions (HU, F, ES) followed the same sense. Thus, three variables (T15, T53 and T57) did not follow the gradient of expression. 2) Interdistrict correlations : Interdependence of a variable expressing beyond a dental district was tested. The objective was to identify whether the effect of the field could also be demonstrated on several dental classes. The same procedure for the decision was used and a single variable (T16) was considered statistically significant. 3) Intertrait correlations : The association between variables expressed on opposed arcades was measured, being the relationship between Carabelli’s trait (T24) observed on upper molars and the protostylid (T54) on the lower molars. We selected this since it seemed that this would be the correlation most often encountered in the literature. A single association was positive, being Carabelli’s trait on the third upper molar with the expression of the protostylid on the second lower molar. • Sexual dimorphism The final point focuses on the measurement of sexual dimorphism in the expression of non-metric dental traits. The analysis was done for only two regions (CZ and CH) given the limited number of samples for which sexual determination could be made. To claim that a biological significance is present, differences in the expression of a variable should present an interpopulational aptitude. We have thus chosen to retain this association as relevant only if it is demonstrated significant for the two populations tested. However, none of the variables was significant. There was no preliminary treatment for standard dental data. Depending on the data and the results of different tests, certain decisions were made. Based on the analyses, it would be perhaps useful and possible to pay attention to certain results of preliminary treatment in order to integrate or exclude certain variables. Chapter 7 This chapter presents two aspects of data analysis. The first focuses on the presentation of the procedure followed for preliminary statistical treatment of the data. The second presents the analyses themselves. • The procedure followed 1) It was decided to work with population frequencies. We have thus calculated the incidences for each variable within each assemblage tested. These are dichotomous or graduated variables, and so the method of expression count developed by Turner (1985) was used, which obtained an adjusted frequency taking into account all of the information provided by the gradations. It was carried out in two steps : obtainment 24 Abstract of a unique frequency followed by the introduction of a correction factor taking into account the sample size. 2) We then selected variables that could be used in the analyses. During preliminary data treatment, we saw that traits expressed within a dental class were not independent. We thus retained one tooth per dental district that we termed the key tooth for all of the variables, including traits that did not seem to follow the gradient of expression (T15, T53 and T57). The dental tuberculum (T16) presents an interdependence between dental classes, which is why we chose only a single tooth on which this trait is expressed, generally the canine. After preliminary statistical treatment of the data, 79 variables were deemed suitable for analysis. 3) We retained several types of multivariate analyses : ascendant hierarchical classification (using Ward’s algorithm) complemented by bootstrap analysis, multidimensional scaling complemented by minimal spanning tree and principal components analysis. • Analysis of the assemblages Several different aspects were addressed. For each domain studied (eastern, southern and articulation [Switzerland]), an individual analysis for each region was carried out, followed by an analysis for each domain. Switzerland was first compared with the eastern domain and then with the southern domain. We then continued analyses with a study including only Bell Beaker samples for the five regions treated. We concluded the analytical phase with a study of the Czech corpus with the sexes separated. What elements were retained? The eastern domain Analyses for Bohemia were based on 13 assemblages (4 Corded Ware, 4 Bell Beaker and 5 Unetice) and 54 variables. The different analyses support the idea of more heterogeneous populations during the Final Eneolithic in Bohemia, which likely reflects higher degrees of mobility or population exchange during these periods. The beginning of the Early Bronze Age, by contrast, reveals a strong cohesion of the groups studied, suggesting a certain degree of isolation from external influences. The intermediary position of the Corded Ware culture supports its involvement in the establishment of the Bell Beaker and the Unetice cultures. The analyses for Hungary were made for four assemblages (Bell Beaker, Obedà-Pitvaros, Perjamos and Nagyrev) and 35 variables. The Nagyrev culture seems to have a strong central position in relationships between the different cultures studied. The Bell Beaker is separated from the other groups. It is neither close to the southern culture with which it is contemporaneous (Obedà-Pitvaros), nor to the Nagyrev with which it is in part contemporaneous - in its early phase - and which succeeds it. Analyses of the eastern domain are based on 17 assemblages (for Bohemia the four Corded Ware, 4 Bell Beaker and 5 Unetice, four groups for Hungary) and 30 variables. For Bohemia, two groupings (Unetice/eastern Corded Ware and Corded Ware/Bell Beaker) were distinguished. The position of the Hungarian groups within this configuration is quite interesting since the Bell Beaker is moderately associated with the first group and the three other assemblages - Nagyrev, Obedà-Pitvaros and Perjamos in the second group. The southern domain Analyses for southern France were based on seven assemblages (two Final Neolithic, two Chalcolithic, one Bell Beaker, one Chalcolithic/Bell Beaker and one Chalcolithic/Early Bronze Age) and 57 variables. Sites for the Final Neolithic and the Chalcolithic show a more heterogeneous situation. By contrast, more recent sites are isolated. Analyses for northern Spain were based on seven assemblages (three Final Neolithic, two Chalcolithic, one Bell Beaker and one Final Neolithic/Bell Beaker) and 17 variables. The different samples for the Final Neolithic are fairly uniform. The more recent assemblages, by contrast, seem to show greater variability. The position of the Bell Beaker complex intermediary between the populations of the Final Neolithic and the Chalcolithic is quite interesting. 25 Analyses of the southern domain were carried out on 13 assemblages (seven French and six Spanish) and 30 variables. The situation shown for this domain is interesting since we find more marked differences for the earlier periods and a certain degree of cohesion for the more recent populations, probably reflecting more marked exchanges between them. The Bell Beaker complex within this configuration lies between these two currents. At the articulation between the two domains Analyses for Switzerland were based on 10 assemblages (five Middle Neolithic groups, two Recent Neolithic, two Bell Beaker and one Early Bronze Age) and 49 variables. The different analyses are in agreement to support the idea of a certain harmony in Middle Neolithic populations and variability in later populations, likely reflecting mobility or a moderate population contribution beginning at the end of the Final Neolithic. Analyses for the relationships between Switzerland and the eastern domain were based on 22 assemblages (five Swiss groups, 13 Czech and four Hungarian) and 30 variables. We observe the same tendencies distinguished during regional analyses. The Swiss sites do not mix with the eastern domain and the Hungarian Bell Beakers seem much closer to their contemporaneous western groups. Analyses of the relationships between Switzerland and the southern domain were based on 18 assemblages (five Swiss groups, seven French and six Spanish) and 30 variables. The Swiss samples are strongly grouped in the assemblage of configurations. They are associated with recent Iberian and French periods. Early groups of the southern domain present higher variability, probably due to mobility or minor population exchanges during these periods. This does not appear to be the case in Switzerland and for the later southern groups, where the apparent homogeneity accentuates external influences. Relationships between Bell Beaker populations This analysis was based on 11 assemblages (four Czech groups, one Hungarian, two Swiss, two French and two Spanish) and 30 variables. A clear dichotomy between eastern and southern Bell Beaker populations is demonstrated by the analyses. The Swiss Bell Beaker groups are clearly associated with the southern domain. Exchanges and external influences seem to be major in the western area of the Bell Beaker complex. Bohemia : analysis with sexes separated The final analysis comprises six groups (men and women, Corded Ware, Bell Beaker and Unetice) and 44 variables. Men in the three cultures are close and women have a tendency to be more variable. Results as easily assimilated within the regional study : a Unetice group, greater variability for Corded Ware and Bell Beaker assemblages and a position of Corded Ware men as a common denominator uniting the group of populations. Chapter 8 The five occupations analyzed offer a rich and complex portrait of society during the 3rd millennium BC. A single explanation does not account for the entire range of variability in the Bell Beaker phenomenon. The involvement of local populations in the emergence of the Bell Beaker varies according to region. It is only in northern Spain and Bohemia that strict links between the Bell Beaker occupation and local occupations exist. For the other three regions, external population influences played a role in the origin of the Bell Beaker occupation, but their contribution also varies. Complete population renewal - or at least a highly significant exogenous impact - is unambiguous for southern France and Hungary. In effect, Bell Beaker populations are clearly distinguished from local populations in these two regions. As for Switzerland, however, shows a local regional population disturbed by partial population renewal or the integration of exogenous individuals. Such a situation in the Swiss region seems to have already been present, although less intensively, during the Late Neolithic. 26 Abstract Dental morphology has demonstrated two populational spheres present in the territory studied. The two entities are clearly distinct; we thus have the southern Bell Beakers and the eastern Bell Beakers, which we respectively term Bell Beakers and Beakers. The southern Bell Beakers are quite similar and the Swiss populations can be strongly linked to their morphology. They form a highly uniform group. The eastern Bell Beakers show a certain cohesion that seems, however, to have been less isolated. Based on these results, it is possible to propose a diffusion model for the Bell Beaker phenomenon : The emergence of the Bell Beaker culture in the southern sphere resulted from the displacement of individuals from the Iberian Peninsula into Europe. The biological impact was recorded to at least Switzerland, and possibly also to Hungary. Thus, the Bell Beakers - small groups of individuals equipped with their material culture and know-how - formed the basis for Bell Beaker diffusion in this region of the phenomenon. The situation in the eastern sphere is more complex. Dental data suggest evolution within a single society. Nevertheless, women - Corded Ware and Bell Beaker - were differentiated from the local populations, probably resulting from societies practicing exogamy. For N. Brodie (2001), it was eastern Bell Beaker women who integrated the western region. Our data support neither this hypothesis nor the inverse. Dental morphologies strongly diverge in the two spheres and the population basis of Bell Beaker women was not part of the southern zones studied here. Thus, to understand the modalities for the establishment of the Bell Beaker phenomenon, we must dissociate the diffusion of southern elements from the exogamic diffusion of women in the eastern domain into two distinct points in times. On the basis of currently available radiocarbon dates suggesting a southwest-northeast gradient for the expansion of the Bell Beaker, we propose the following : 1. Migration of groups of Bell Beaker individuals from the Iberian Peninsula toward the east, while the eastern domain is still occupied by the Corded Ware culture. 2. Part of the Corded Ware on the edge of the phenomenon was individualized and adopted, by borrowing, some of the southern Bell Beaker traditions. Diffusion of this new society - the Beakers continued toward the east. At the same time, certain eastern elements were diffused toward the west. 27 Introduction Et bien oui, encore une thèse abordant le Campaniforme! Intriguant... irritant... le Campaniforme, précédé, indifféremment, par énigme, phénomène ou encore question, a fait l’objet de centaines d’écrits qui l’ont examiné sous toutes ses coutures. Nous pourrions croire que tout a déjà été pensé et repensé, dit et redit, écrit et réécrit, et pourtant, pour citer l’une des spécialistes de cette entité de la fin du Néolithique1, plus nous travaillons sur le Campaniforme, moins nous le comprenons! Quel défi - ou plutôt, quelle douce folie - imaginer fournir des éléments de compréhension inédits. Pour certains chercheurs, le Campaniforme ne désigne qu’une céramique - décorée et en forme de cloche renversée - largement répandue sur l'ensemble de l'Europe pendant le 3e millénaire avant notre ère. Cela est vrai, mais en partie seulement, puisque le Campaniforme constitue une entité qui se singularise des autres manifestations archéologiques, non seulement par son extension géographique particulièrement vaste, mais également par la complexité évidente de ses vestiges. Que savons-nous aujourd’hui du Campaniforme ? Il se développe à l’échelle de l’Europe suivant un gradient sud-ouest - nord-est lors du 3e millénaire avant notre ère sur des substrats locaux très différents. Il est constitué par une culture matérielle fédératrice (céramique décorée, céramique commune, brassards d’archer, poignards en cuivre, pointes de Palmela, boutons en os perforés en V...) et par des structures funéraires (sépulture individuelle ou collective, inhumation ou incinération) et domestiques (construction sur poteaux ou en pierres sèches, de forme circulaire, ovalaire, rectangulaire...) variées. Les premières interprétations à propos de l'apparition du Campaniforme se sont fondées sur le caractère paneuropéen de la céramique décorée notamment. Cette spécificité a engendré une multitude de scénarii des plus colorés, complémentaires ou non, naviguant entre mouvements de populations, circulations des biens ou encore des idées. De nos jours, les différences et les éléments moins exceptionnels de la culture matérielle (céramique commune ou industrie lithique) sont à l’honneur et distinguent une certaine opposition des aires occidentale et orientale du Campaniforme. Pour comprendre les mécanismes liés à l’émergence du Campaniforme, il est nécessaire de travailler sur un vaste territoire puisqu’il s’agit d’un phénomène étendu à l’ensemble de l’Europe. Et c’est dans cette perspective de vision large du Campaniforme qu’un programme de recherche interdisciplinaire, intitulé L'Europe du 3e millénaire avant notre ère : des faits archéologiques à l'interprétation du peuplement2, a vu le jour. Ce programme vise une meilleure compréhension des modalités d’apparition du Campaniforme en évaluant l’importance du fonds néolithique dans sa mise en place. Il comporte quatre volets complémentaires - typologie et chronologie de la céramique commune, occupation du territoire, métallurgie et anthropologie biologique. Ainsi notre travail constitue le dernier volet : étudier les individus qui sont à la source du Campaniforme. Témoins privilégiés d’une culture que l’on a encore du mal à définir dans son entièreté, ils peuvent fournir des clés de compréhension intéressantes. Cependant, s’intéresser aux individus qui sont à l’origine du Campaniforme n’est pas un élément nouveau. Plusieurs auteurs se sont attelés à la tâche, de l’étude ponctuelle à l’échelle européenne, de l’observation de la forme du crâne à la composition chimique des ossements. Et la mobilité est un terme profondément lié à l’émergence du Campaniforme à partir du moment où l’individu y est engagé. Elle est caractérisée de partielle, totale ou différentielle. La morphologie crânienne particulière des individus campaniformes en est la cause. En effet, l’individu campaniforme est extrêmement brachycrâne (crâne arrondi), à aplatissement occipital et totalement 1 C’est à la professeure Marie Besse que l’on doit ces mots. Programme de recherche (2003-2006) dirigé par la professeure Marie Besse et financé par le Fonds national de la recherche scientifique suisse (FNS subside 100012-100599). 2 29 différent du substrat local (Menk 1979, p. 272). On a longtemps considéré cette morphologie comme un indice irréfutable de mouvements de populations à l’origine du Campaniforme. Mais le va-et-vient des interprétations issues exclusivement des composantes matérielles a amoindri l'intérêt de l’individu, et dès lors de son étude. Notre recherche intervient à nouveau sur l’individu campaniforme en l’abordant par l’étude de sa dentition, et plus précisément par l’analyse des traits non métriques. Ce sont des variations anatomiques observées sur l’ensemble de la dentition permanente et déciduale. On se réfère, en général, à des caractères qui sont présents ou absents, ou qui manifestent différents stades de développement. Ces caractères présentent l’avantage, et non des moindres, de pouvoir être observés sur le vivant permettant ainsi d’avoir une meilleure connaissance de leur valeur biologique en tant qu’indicateurs des distances biologiques entre les populations. La dent possède également l’atout d’être l’une des parties la plus résistante du squelette et par conséquent représente souvent l’élément le mieux conservé en milieu archéologique. Nous avons choisi pour cette étude cinq régions représentant cinq portraits différents de l’occupation campaniforme pendant le 3e millénaire avant notre ère. Deux régions appartiennent au domaine oriental : la Bohême - territoire considéré comme un acteur important dans l’histoire du peuplement campaniforme, représente l’occupation orientale type - et la Hongrie - son occupation campaniforme est seule dans l’aire orientale hors substrat cordé et se limite à la région bordant Budapest. Deux régions appartiennent au domaine méridional : le sud de la France - les données archéologiques révèlent généralement deux phases d’occupation campaniforme, la première marquant une rupture avec les traditions locales et la seconde révélant une période d’intégration et de cohabitation - et le nord de l’Espagne - région importante dans la compréhension du phénomène Campaniforme, il s’y développe dans une phase récente du Chalcolithique et coexiste avec lui jusqu’à l’émergence du Bronze ancien. La dernière région sélectionnée est la Suisse située en position géographique intermédiaire, elle laisse apparaître la dualité des traditions des différentes aires culturelles est-ouest. L’objectif est de définir, pour chacune de ces régions, si l’émergence du Campaniforme coïncide ou non avec un renouvellement, même partiel, de la population. Il est également intéressant d’appréhender les relations entre les différents groupes du Campaniforme sur l’ensemble du territoire étudié afin de proposer un scénario de dynamique du peuplement pendant le 3e millénaire avant notre ère. Notre travail s’articule en huit parties : Le chapitre 1 approche l’essence même de ce travail : le Campaniforme. Cette présentation n’a pas la prétention d’être exhaustive, elle offre toutefois un portrait synthétique des multiples facettes de ce complexe. Nous évoquons ce qui caractérise cette entité sur un territoire aussi vaste, les interprétations d’hier et d’aujourd’hui. Nous situons également notre recherche et ses objectifs. Le chapitre 2 aborde la description des cinq régions intégrées dans notre recherche. Nous rappelons l’insertion culturelle, spatiale et temporelle du Campaniforme pour chacune d’entre elles. Nous présentons également les données provenant de l’archéologie et de l’anthropologie biologique ainsi que les divers échantillons analysés. Le chapitre 3 est un lexique des termes spécifiques à notre recherche sous forme de texte pour en faciliter la lecture. Le chapitre 4 a pour objectif de montrer que la morphologie dentaire peut se révéler un outil intéressant dans les analyses concernant la compréhension des relations entre populations. D’abord, nous nous intéressons à la valeur biologique de ces variables (développement de la dentition, mode de transmission, influences environnementales, héritabilité...). Ensuite, nous nous attachons à la fiabilité du système d’observation. Enfin, nous contrôlons si en application sur des populations actuelles ces variables s’avèrent suffisamment sensibles pour être considérées comme de bons estimateurs des distances biologiques entre populations. 30 Introduction Le chapitre 5 concerne la sélection des variables non métriques dentaires, le mode d’acquisition et l’évaluation de la concordance des données (déterminations dentaires et observation des traits non métriques). Le chapitre 6 aborde le traitement préliminaire des observations dentaires. Il comporte divers aspects : une phase descriptive servant à vérifier la transcription des données et à écarter les données non pertinentes, un traitement des observations bilatérales en données uniques afin d’éliminer les informations redondantes fournies par les antimères, une évaluation des relations entre les variables (corrélations intradistrict, interdistrict et intertrait) et, enfin, la mesure de l’influence du sexe dans l’expression des traits non métriques dentaires. Le chapitre 7 présente l’analyse des données. Il comporte deux volets : le premier touche la procédure suivie lors du traitement statistique des données, le second s’intéresse aux analyses proprement dites. Une discussion des résultats obtenus est réalisée dans le chapitre 8. Nous présentons, sur la base des éléments fournis par l’analyse des traits non métriques dentaires, un modèle de peuplement et déterminons, pour chacune des régions étudiées, si un renouvellement de la population a eu lieu ou non avec l’apparition du Campaniforme. Enfin, nous proposons une synthèse du Campaniforme à l’échelle du phénomène et définissons les perspectives d’études et les nouvelles questions. 31 Chapitre 1 Le Campaniforme... un phénomène... une énigme Le Campaniforme se réfère initialement à un style de céramique largement répandu en Europe et en Afrique du Nord pendant le 3e millénaire avant notre ère, période qui correspond à la fin du Néolithique. Il a été défini pour la première fois à la fin du 19e - début du 20e siècle, en Espagne, pour décrire des vases à profil en S, en forme de cloche renversée. Le Campaniforme touche un territoire vaste allant des îles britanniques aux côtes d’Afrique du Nord - pour son extension nord-sud - et du Portugal à la Hongrie - pour son extension ouest-est. Nous n’avons pas la prétention de faire un bilan complet de l’histoire des recherches sur le Campaniforme, certains s’y sont déjà attelés et l’ont très bien réalisé. Nous pouvons citer, à titre d’exemple, les synthèses d’A. Gallay (1988, 2001), de C. Strahm (1998), de M. Bailly (2002), L. Salanova (2005) ou encore de M. Vander Linden (2006). Il s’agit pour nous d’évoquer les faits, de présenter rapidement ce qui a été dit et, surtout, ce qui est dit aujourd’hui, car les recherches sur le Campaniforme s’étendent sur plus d’un siècle ! 1.1. La situation précédant le Campaniforme Nous allons nous intéresser à la situation précédant le Campaniforme en survolant les différents contextes culturels sur lesquels il s’est développé. Nous préciserons les contextes d’apparition du Campaniforme des régions intégrées dans ce travail lors du prochain chapitre (chapitre 2). Le plus important groupe culturel sur lequel le Campaniforme se met en place est le Cordé. Il s’étend sur l’Europe septentrionale et une grande partie de l’Europe centrale. On différencie plusieurs ensembles régionaux au sein du Cordé, comme le groupe de Mansfeld (Allemagne centrale), celui des sépultures individuelles (Danemark, Pays-Bas, nord de l’Allemagne), le Lubaczow (sud-est de la Pologne), l’Auvernier-Cordé (est de la France, ouest de la Suisse)... (Vander Linden 2006). Cette culture est davantage connue par son rituel funéraire aux règles d’inhumation uniformes (orientation des corps et matériel d’accompagnement) et se distingue par son absence de structures d’habitats à l’exception de la Suisse où des habitats palafittiques sont biens connus. Ces éléments conduisent à considérer les cordés comme des nomades parcourant l’Europe. De l’origine locale, avec différents foyers d’apparition potentiels telles la culture des amphores globulaires (est de l’Allemagne, Pologne, Russie), celle des gobelets à entonnoir (nord de la plaine européenne et Scandinavie) ou encore la culture de Mansfeld (nord de l’Allemagne), à la migration de populations depuis les steppes pontiques, la genèse du Cordé, comme celle du Campaniforme, anime les discussions. Seul le Campaniforme hongrois appelé le groupe Campaniforme-Csepel apparaît, dans la partie orientale, hors contexte cordé. Le substrat sur lequel il s’installe, est le groupe de Makó - stade initial du Bronze ancien - (Endrödi 1998). Ainsi, contrairement au reste de l’Europe précampaniforme, qui se situe dans une phase finale du Néolithique, en Hongrie le Campaniforme se développe au sein du Bronze ancien. L’unité précampaniforme orientale ne se retrouve pas en Europe occidentale. En effet, cette dernière se caractérise par une série de groupes culturels distincts et limités dans l’espace attribué au Néolithique récent, à l’Enéolithique ou encore au Chalcolithique selon les régions. Cette hétérogénéité n’exclut pas d’éventuels contacts et apparentements entre les cultures de la fin du Néolithique occidental (Vander Linden 2006). Le Campaniforme se construit ainsi sur ces substrats variés et cohabite, en partie, avec certains d’entre eux. En France, pour ne citer qu’un exemple, le Campaniforme se développe sur - et en parallèle à - un certain nombre de groupes culturels, comme l’Artenac, le Fontbouisse, le Gord-DeûleEscaut ou encore le Vérazien... (Guilaine 1998a). 33 Chapitre 1 1.2. Les faits Nous survolons ici les différents éléments définissant le Campaniforme. Nous aborderons dans un premier temps le matériel qui le caractérise. Ensuite nous nous intéresserons à la diversité des structures liées à ce complexe. Finalement, nous situerons chronologiquement ce phénomène. 1.2.1. Le matériel attribué au Campaniforme Le Campaniforme est défini au départ par un type de céramique en forme de cloche renversée retrouvée principalement en contexte funéraire (figure 1). Celle-ci est richement décorée par des motifs géométriques simples, notamment par des bandes hachurées, des lignes horizontales, des triangles ou encore des chevrons (Salanova 2000). On peut distinguer parmi ce type de production deux catégories. La première est nommée par L. Salanova (2000, 2002) le standard. Ce type de céramique possède surtout une coloration orangée, les décors sont standardisés par des lignes horizontales et des bandes hachurées. Le tracé des décors est fin et régulier souvent obtenu par impression de coquillages (Salanova 2002). Ces céramiques ont une large répartition géographique. La seconde catégorie regroupe des productions régionales. Ce type de céramique présente une facture moins soignée que le standard. Sa teinte est en général brune et son décor réalisé par incision et impression au peigne (Salanova 2002). Figure 1 : La céramique décorée campaniforme A gauche, un vase décoré provenant du site de Plisting bei Landau an der Isar (Allemagne). D’après B. Engelhardt 1999, p. 38. A droite, un gobelet décoré provenant de l’hypogée de Marinaru en Sardaigne (Italie). D’après E. Contu, 1998, p. 286. Le Campaniforme est également constitué d’une céramique domestique (figure 2) que l’on appelle céramique commune, lisse, d’accompagnement voire, aujourd’hui, de complémentaire (Strahm 2004). Elle est peu ou pas décorée et sa réalisation est moins soignée (Besse 2001). Elle présente une grande variabilité morphologique, dimensionnelle et de composition des pâtes (Besse 1998a). Elle est constituée de pichets, d’écuelles, de vases, de jarres et de cuillères pour ne citer que quelques exemples. Cette céramique se retrouve essentiellement en contexte d’habitat et, contrairement à celle décorée, varie en fonction des régions et des traditions locales (Besse 2003, 2004a). Figure 2 : La campaniforme céramique commune Trois exemplaires bien conservés et de formes diverses provenant du site d’habitat de Alle, Noir-Bois dans le Jura (Suisse). Photo : office du Patrimoine historique du Jura. De petites pièces rectangulaires en pierre présentant des perforations aux extrémités sont interprétées comme des brassards d’archer (figure 3). Si la fonction de protection du bras lors du tir à l’arc a souvent été discutée en raison de la petitesse de certaines pièces, l’analyse de l’emplacement de celles-ci dans les 34 Le Campaniforme... un phénomène... une énigme sépultures individuelles confirme une position au niveau de l’avant-bras (Besse 2006, p. 37). On les retrouve en contexte funéraire, et de préférence - mais non de manière exclusive - dans les tombes masculines. Des petits poignards en cuivre à lame triangulaire lisse et à languette d’emmanchement (figure 3) et des armatures de flèches perçantes triangulaires et à retouche bifaciale couvrante font également partie du mobilier caractérisant ce complexe (Bailly 2002). Ce matériel est en général mis au jour en contexte funéraire. Figure 3 : Le matériel associé au Campaniforme A gauche, un brassard d’archer provenant de l’hypogée de Fosso Conicchio à Viterbo (Italie). D’après F. Nicolis et E. Mottes 1998, p. 199. A droite, une lame de poignard a été retrouvée dans le dolmen de Saint-Eugène dans l’Aude (France). D’après J. Guilaine 1998b, p. 107. Le Campaniforme possède également divers éléments caractéristiques même si ces objets ne se retrouvent pas sur l’ensemble de l’aire touchée par ce complexe. Il s’agit de pointes en cuivre dites de Palmela, des pendeloques arciformes, des parures en or et en argent, des boutons en os perforés en V (figure 4). Figure 4 : Le matériel associé au Campaniforme A gauche, une pointe de Palmela provenant des mines de Cabrières dans l’Hérault (France). D’après J. Guilaine 1998b, p. 104. A droite, des boutons en os perforés en V provenant de la tombe collective de Padre Jossu à Cagliari (Italie). D’après F. Nicolis et E. Mottes 1998, p. 323. On ne peut terminer la caractérisation du Campaniforme sans parler de sa riche iconographie prenant la forme de stèles anthropomorphes (figure 5). Ces magnifiques monuments richement parés présentent des ornementations comparables à celles des gobelets décorés. Figure 5 : Le matériel associé au Campaniforme Stèle anthropomorphe campaniforme provenant du site du Petit-Chasseur à Sion (Suisse). D’après S. Favre et al. 1986, planche 21. Voici en quelques mots, un survol des principaux éléments de la culture matérielle permettant de définir le Campaniforme. 35 Chapitre 1 1.2.2. Les structures funéraires Les structures funéraires campaniformes montrent une grande diversité sous une apparente homogénéité régionale. La dichotomie, généralement adoptée, pour synthétiser le rituel funéraire campaniforme annonçant des sépultures individuelles dans l’aire orientale du phénomène et des sépultures collectives dans l’aire occidentale n’est pas incorrecte mais restreint fortement la variabilité existante. En effet, on passe de la sépulture individuelle en fosse, ou en coffre, à la sépulture individuelle à l’intérieur même d’une sépulture collective, et à la construction de monuments mégalithiques, voire à l’incinération (figure 6). • Le domaine oriental Dans le domaine oriental, le rituel funéraire en usage presque exclusif pendant le Campaniforme est l’inhumation individuelle en pleine terre. Il est tout de même possible de mettre en évidence, selon les régions, des sépultures collectives, des sépultures dans des coffres en bois, en cistes et des incinérations (Harrison 1986). Ces dernières, en contexte campaniforme, suivent un axe est-ouest avec une fréquence atteignant 90% en Hongrie et 3% en Allemagne (Müller 1998). La Tchéquie est une région d’une grande importance ; la Bohême et la Moravie regroupent à elles seules environ 2000 tombes campaniformes qui sont respectivement plus nombreuses pour la première et plus riches et variées pour la seconde (Harrison 1986). On retrouve en Bohême un certain nombre de nécropoles comptant en général une vingtaine de sépultures. Le rituel semble homogène sur l’ensemble de la région, avec des inhumations en tombes plates dont l’orientation et l’aménagement des corps et des structures sont constants. Certains sites offrent des tombes cordées et campaniformes, comme à Čachovice (Mlada Boleslav) et à Brandýsek (Slany) (Neustupný et Smrž 1989). La Moravie, quant à elle, compte à la fois des sépultures isolées ou en nécropole. Les cimetières peuvent contenir jusqu’à près de quatre-vingts tombes, tel celui de Holásky II (Brno) ou encore celui de Pavlov (Breclav) (Dvořák 1991). Parallèlement, on assiste à l’apparition de tombes sous tumulus, comme à Smolin (Breclav), phénomène qui se limite au nord-ouest de la région (Dvořák et al. 1996), et à l’apparition de la crémation, comme dans le site de Lechovice (Znojmo) (Medunová et Ondraček 1969). En Autriche, en Slovaquie et en Pologne, les sépultures campaniformes suivent la tradition orientale de l’inhumation individuelle en pleine terre. On peut citer, à titre d’exemple, les sites de Ragelsdorf (Pernersdorf, Autriche) (Neugebauer-Maresch 1994), de Skalica (Senica, Slovaquie) (Vladař 1969) ou encore de Samborzec (Sandomierz, Pologne) (Machnik 1984). L’incinération est également présente, comme à Laa an der Thaya "Bonteufeln" (Mistelbach, Autriche) ou encore à Zwingendorf "Am Kugelzipf" (Mistelbach, Autriche) (Neugebauer-Maresch 1994). En Hongrie, les sépultures campaniformes sont représentées tant par des incinérations que par des inhumations, situation mixte que l’on doit probablement à la culture de Mako, culture précédant le Campaniforme et pratiquant l’incinération. Cette mixité des pratiques se retrouve dans les nécropoles de Bekasmegyer (Budapest) (Kalicz-Schreiber et Kalicz 2001), de Tököl II (Budapest) (Kalicz-Schreiber 1981), de Budakalasz (Budapest) (Neustupný 1984) ou encore de Bucem (Csepel) (Harrison 1980). • Le domaine septentrional Le domaine septentrional montre une variabilité plus importante, de la sépulture individuelle à la réoccupation de monuments mégalithiques. En Angleterre et en Irlande, l’inhumation classique est la sépulture individuelle. Celle-ci est souvent en pleine terre recouverte d’un petit tertre. On a généralement affaire à de petites nécropoles avec au plus une vingtaine de sépultures comme à Eynsham (Oxford) ou à Cassington (Oxford) (Harrison 1986). Aux Pays-Bas, les inhumations sont individuelles et recouvertes d’un petit tertre de sable comme à Exloo (Drenthe) et à Otterberg II (Bronstijd) (Lanting 1973). A Molenaarsgraaf (Zuid-Holland), plusieurs sépultures ont également été retrouvées autour des structures d’habitats (Harrison 1986). En Allemagne, le Campaniforme est principalement attesté par des sépultures individuelles. Celles-ci peuvent être sous tumulus comme à Armsen (Kirchlinteln), à Neuenbülstedt-Wüllenheide (Bülstedt) ou encore à Hünenberg (Uthlede) (Strahl 1990). La région du Mecklembourg-Poméranie occidentale et le 36 Le Campaniforme... un phénomène... une énigme Brandebourg se caractérisent par une réoccupation des sépultures mégalithiques construites pendant la Civilisation des gobelets en entonnoir, ainsi que par des sépultures individuelles en fosse. En Belgique, la documentation est particulièrement hétérogène. On peut tout de même citer les sépultures individuelles sous tumulus de Bergeijkse Heide (Anvers) et les incinérations en fosse de Wijkhuis (Flandre orientale) (Warmenbol 1996). La réutilisation des tombes mégalithiques est également présente avec des inhumations secondaires à l’intérieur des monuments comme dans l’ouest de la France à Villes-es-Nouaux (Jersey), à Kerbors (Côtes d’Armors) (L’Helgouach 1998), à Bréhuidic à Sarzeau (Morbihan), au Danemark à Bigum (Tjele) ou encore à Kirke Helsinge (Gørlev) (Liversage 2003). Figure 6 : Variabilité des structures funéraires à travers l’Europe. D’après M. Besse et J. Desideri 2005, p. 83. 1. Straubing, Bavière, Allemagne (Engelhardt 1999) - 2. Franzhausen, Schottergrube der Firma Traunfellner, Autriche (Neugebauer et Gattringer 1982) - 3. Gemeinlebarn, St Pölten, Autriche (Neugebauer et Neugebauer-Maresch 2001) - 4. Sulejovice, Litoměřice, Bohême, Tchéquie (Moucha 1958) - 5. Pavlov, Breclav, Moravie, Tchéquie (Dvořák 1991) - 6. Samborzec, Sandomierz, Pologne (Nosek 1964) - 7. Samborzec, Sandomierz, Pologne (Makarowicz 2003) - 8, 9, 10. Bekasmegyer, Budapest, Hongrie (Kalicz-Schreiber et Kalicz 2001) - 11. Scharfstädt, Saxe-Thuringe, Allemagne (Otto 1950) - 12. Landau, Süd-Ost, Allemagne (Husty 2004) - 13. Eisleben, SaxeAnhalt, Allemagne (Stahlhofen 1986) - 14. Molenaarsgraaf, Zuid-Holland, Pays-Bas (Louwe Kooijmanns 1974) - 15. Villes-es-Nouaux, Jersey, France (Salanova 2003) - 16. Amesbury, Wiltshire, Grande Bretagne (www.wessexarch.co.uk) - 17. Knowth, Meath, Grande Bretagne (Eogan 1976) - 18. Barnack, Grande Bretagne (Strahm 1995) - 19. Petit-Chasseur, Sion, Suisse (Gallay 1976) - 20. SaintMartin-de-Corléans, Aoste, Italie (Mezzena 1995) - 21. Via Bruschi, Sesto Fiorentino, Italie (Sarti et Martini 1998) - 22. Paradis-auxAnes, Jard-sur-Mer, Vendée, France (Mahieu 1992a) - 23. Champs-Galottes, Yonne, France (Mahieu 1992a) - 24. Les Fados, Pépieux, Aude, France (Mahieu 1992a) - 25. Fontvieille, Bouches-du-Rhône, France (Mahieu 1992a) - 26. La Fare, Forcalquier, Alpes-de-HauteProvence (Lemercier 1997-1998) - 27. Arenero de Soto, Getafe, Madrid, Espagne (Garrido-Pena 2000) - 28. La Vaquera, Segovia, Espagne (Garrido-Pena 2000) - 29. Los Millares, Almeira, Espagne (Strahm 1995) - 30. Canada de Carrascal, Sevilla, Espagne (Leisner et Leisner 1943) - 31. Tres Montes, Navarra, Espagne (Andres et al. 2002) - 32. La Sima, Soria, Espagne (Rojo Guerra et al. 2003) - 33. Los Maioles, Rubiò, Espagne (Clop 2004) - 34. Agualva, Estremadura, Portugal (Tavares da Silva et Soares 1998) - 35. Cova da Moura, Estremadura, Portugal (Strahm 1995) - 36. Pedra Branca, Montum, Alentejo, Portugal (Cardoso 2001) - 37. Ferradeira, Faro, Algarve, Portugal (Tavares da Silva et Soares 1998). 37 Chapitre 1 • Le domaine méridional Le domaine méridional offre une palette plus colorée des rituels funéraires. La plupart des sépultures campaniformes provient de tombes mégalithiques. En effet, la réutilisation de sépultures collectives des périodes antérieures (dolmen, hypogée, grotte naturelle...) est la pratique la plus courante. La Suisse se situe géographiquement entre deux grands domaines, méridional et oriental. Dans la partie occidentale de la Suisse, la nécropole mégalithique du Petit-Chasseur à Sion (Valais) est sans aucun doute le gisement le plus important du Campaniforme suisse (Gallay 1976, Favre et Mottet 1995). La partie orientale de la Suisse reste à l’écart des sépultures mégalithiques. Quelques inhumations individuelles campaniformes en pleine terre suivant la tradition rencontrée dans le domaine oriental ont été mises au jour, comme à Riehen (Bâle) ou encore à Allschwill (Bâle) (Moinat et Stöckli 1995). En Italie septentrionale, les sépultures sont relativement rares. Elles sont, en général, individuelles comme dans les sites de Ca di Marco à Gambara (Brescia) (Cornaggia Castiglioni 1971) ou de Roccolo Bresciani (Brescia) (Aspes et al. 1980). On trouve tout de même des sépultures collectives comme à Saint-Martin de Corléans à Aoste (Aoste) (Mezzena 1985) ou encore Riparo Valtenesi à Manerba del Garda (Brescia) (Barfield 1983). En Italie centrale, on note la présence de quelques sépultures, comme le tumulus dans le site de Via Bruschi à Sesto Fiorentino (Florence) (Sarti et Martini 1998) ou encore la Grotta della Scaletta à Presso Vecchiano (Pisa) (Ceccanti et al. 1981). Les connaissances portant sur le Campaniforme en France sont inégalement réparties sur l’ensemble du territoire (Guilaine et al. 1988). Le Campaniforme du centre-ouest se retrouve dans des sépultures collectives comme le dolmen E 136 à Taizé (Deux-Sèvres) (Hébras 1965) ou encore le dolmen sous tumulus de Bernet à Saint Sauveur (Gironde) (Roussot-Larroque 1990). On rencontre également des sépultures individuelles sous tumulus comme au Paradis-aux-Anes à Jard-sur-Mer (Vendée) (Mahieu 1992a). Les sépultures du Bassin parisien et du nord de la France sont soit des sépultures individuelles comme à Jablines (Seine-et-Marne) ou encore à Champs-Galottes (Yonne), sous tumulus ou non, soit attestées à l’intérieur même des sépultures collectives mégalithiques construites antérieurement au Campaniforme, d’origine Seine-Oise-Marne, comme à Epone (Yvelines) (Constantin et Blanchet 1998). Le Campaniforme du sud de la France est documenté par un nombre important de sites et tous les types de sépultures campaniformes y sont présents (Mahieu 1992a). Ce sont en majorité des sépultures collectives, construites par des groupes culturels antérieurs et réutilisées. Les sépultures collectives peuvent être architecturalement variées, avec des sépultures mégalithiques (dolmens), des hypogées ou des grottes sépulcrales. On peut citer notamment le dolmen des Fados à Pépieux (Aude) (Mahieu 1992a), l’allée couverte de Jappeloup à Trausse (Aude) (Guilaine 1967), l’hypogée de Fontvieille (Bouches-duRhône) (Mahieu 1992a), la sépulture en grotte du Cimetière à Châteaurenard (Bouches-du-Rhône) (Courtin 1974) ou encore la grotte Murée de Montpezat (Alpes-de-Haute-Provence) (Mahieu 1992a). Quelques tombes semblent tout de même n’être qu’exclusivement campaniforme comme le tumulus du Gendarme (Var) (Vaquer 1998). Certaines inhumations individuelles aménagées dans des fosses comme le site de la Fare à Forcalquier (Alpes-de-Haute-Provence) ont pu être mises au jour (Lemercier 19971998). En Espagne, les populations campaniformes ont utilisé différents types de structures funéraires. On trouve des tombes ovales individuelles en pleine terre comme à Valdeprados (Avila) (Garrido-Pena 2000). On rencontre également des tombes individuelles en ciste comme à Fuente Palmera (Cordoue) (Martin Socas et al. 1998). L’inhumation en grotte est pratiquée à Balma de Solanells (Lérida), sépulture présentant une trentaine d’individus (Martin Colliga 1998). Les sépultures collectives sont aussi présentes. On peut citer la tombe mégalithique de San Martin de la Guardia (Alava) ou encore l’inhumation multiple d’Agoncillo (Logrono) qui prend la forme d’un grand puits ovale où l’on dénombre plus de 70 individus en position forcée (Harrison 1986). Des nécropoles tumulaires sont présentes en Galicie comme à As Rozas ou à As Pereiras (Prieto Martinez 1998). L’inhumation individuelle au sein d’une sépulture multiple a également été mise en évidence. On peut notamment citer le tumulus de la Sima (Soria) dans lequel trois individus campaniformes ont été inhumés dans le monument construit à une époque antérieure (Rojo Guerra et al. 2003). 38 Le Campaniforme... un phénomène... une énigme Au Portugal, parallèlement à la réutilisation de structures funéraires collectives datant des périodes précédentes, on trouve un nouveau type de sépulture collective, les tholos, des monuments à fausse coupole. On peut citer le site de Agualva (Estremadura) (Tavares da Silva et Soares 1998), dont la structure est constituée d’une chambre circulaire avec un couloir d’accès. Le Campaniforme est également attesté par des sépultures en grotte comme à Cova da Moura (Estremadura) (Tavares da Silva et Soares 1998). 1.2.3. Les structures domestiques Les structures domestiques campaniformes montrent également une grande diversité. Du type de construction - sur poteaux en Europe orientale et centrale et en pierres sèches dans le Bassin méditerranéen -, à la forme des maisons - circulaires, ovalaires, rectangulaires -, ou encore à leur localisation - à proximité ou non des habitats du Néolithique final par exemple -, les sites domestiques du Campaniforme se ressemblent peu (figure 7). • Le domaine oriental Si le rituel funéraire du domaine oriental est relativement bien documenté, la situation n’est pas la même pour les structures domestiques. Bien que la quantité d’habitats ne soit pas insignifiante - plus d’une centaine de sites -, on trouve essentiellement des fosses le plus souvent isolées comme dans le sud de l’Allemagne à Prunn (Eichendorf) ou à Altötting-Kapellplatz (Heyd et al. 2004). La reconstitution de villages, ou tout simplement de maisons, est plus rare. Une habitation a pu être restituée sur la base de l’analyse des trous de poteaux en Hongrie à Szigetszentmiklos-Udülösor (Budapest) présentant un plan allongé, elliptique, avec un toit à double pente (Endrödi 1998). En Tchéquie, les habitats sont d’une grande importance pour la connaissance du Campaniforme oriental, car dans la plupart des cas, il s’agit de fosses détritiques sans autre contexte, et celles-ci renferment des formes céramiques inconnues des tombes (Bertemes et Heyd 1996). • Le domaine septentrional Les habitats campaniformes du domaine septentrional sont connus notamment par la maison I du célèbre site de Molenaarsgraaf aux Pays-Bas (Louwe Kooijmans 1974). La forme de cette maison est allongée aux extrémités légèrement en pointe. En Angleterre et en Irlande, les structures domestiques prennent en général la forme de maisons ovalaires comme à Ballaynagilly (Irlande du Nord), Belle Tout (Sussex) ou encore Gwithian (Cornouailles) (Harrison 1986). La structure d’habitat campaniforme la plus importante au Danemark est le site de Myrhoj (Jutland), dans lequel trois maisons rectangulaires ont pu être mises au jour (Harrison 1986). • Le domaine méridional Le domaine méridional est sans aucun doute celui qui livre le plus d’informations concernant les structures domestiques. Le nombre de maisons, et même de villages, est important. En Suisse, il s’agit principalement de sites terrestres, on dénombre cependant de rares découvertes en contexte lacustre (Besse 1998b). Des deux habitats principaux, seul le gisement de Champ-Vully Est à Rances (Vaud) offre la possibilité de reconstituer des plans d'habitation. Deux cabanes ont été mises au jour dans les vestiges liés à la transition Campaniforme-Bronze ancien et trois autres bâtiments sont rattachés à l'occupation campaniforme (Gallay et Baudais 1985, Besse à paraître). En Italie septentrionale, les sites campaniformes sont principalement des découvertes en contexte d’habitat - en grotte, en abri ou en plein air - dont les surfaces de fouille, souvent petites, ne permettent pas de préciser les plans (Bermond Montanari 1998). En Italie centrale, on rencontre quelques sites d’habitats, comme dans la région de Sesto Fiorentino, et le site de la Querciola (Florence) (Sarti 1998, Sarti et Martini 2001). En France, les habitats de la région du centre-ouest sont peu nombreux. On constate la présence de sites uniquement campaniformes, comme celui de la Grande Pigouille à Belluire (Charente-Maritime) (Bouchet et al. 1990), ou celle de sites où le Campaniforme est associé à des éléments du Néolithique 39 Chapitre 1 final, comme dans le site des Loups à Echiré (Deux-Sèvres) (Burnez et Lagarde 1986). Le Campaniforme du sud de la France est documenté par plusieurs habitats. On remarque la présence d’habitats campaniformes « purs », comme le site de Embusco 3 à Mailhac (Aude) (Taffanel et Taffanel 1957). On peut différencier les habitats campaniformes “purs”, comme celui de Embusco 3 à Mailhac dans l’Aude, de ceux dits « mixtes », soit avec du Campaniforme et des éléments assignés à des groupes culturels autres. On soulignera la présence de sites particuliers : les fossés remplis d’artefacts campaniformes encerclant des bâtiments dont la fonction vraisemblablement très spécifique demande encore à être précisée comme le site de Le Mourral à Trèbes dans l’Aude (Vaquer 1998) ou encore celui de Médor à Ornaisons (Aude) (Guilaine et al. 1989). Dans la région de l’est et du centre-est, deux gisements semblent importants dans l’approche du Campaniforme domestique. Ce sont les habitats de Derrière-le-Château à Géovreissiat et Montréal-la-Cluse (Ain) (Hénon et Vérot-Bourrely 1998) et de La Noue à Saint-Marcel (Saône-et-Loire) (Amrane et Ducreux 1996, Salanova et Ducreux 2005). L’abri de l’Aulp 1 à SaintBernard (Isère) est l’un des rares sites localisé en altitude (1700m), il est le témoin d’une occupation vraisemblablement brève et ponctuelle, attestant des passages intra-alpins par les cols secondaires (Morin 1998). Figure 7 : Variabilité des structures domestiques à travers l’Europe. D’après M. Besse et J. Desideri 2005, p. 84. 1. Altötting-Kapellplatz, Allemagne (Heyd 2004) - 2. Kozly, Bohême, Tchéquie (Husty 2004) - 3. Horní Lán, Olomouc-Slavonín, Moravie, Tchéquie (Turek et al. 2003) - 4. Žadovice, Moravie, Tchéquie (Turek et Peška 2001) - 5. Smarglin, Kujawy, Pologne (Czebreszuk 2003) - 6. Szigetszentmiklós-Üdülösor, Budapest, Hongrie (Husty 2004) - 7. Mirhoj, Jutland, Danemark (Harrison 1986) 8. Molenaarsgraaf, Zuid-Holland, Pays-Bas (Louwe Kooijmanns 1974) - 9. Gwithian, Cornouailles, Grande Bretagne (Harrison 1986) 10. Belle-Tout, Sussex, Grande Bretagne (Harrison 1986) - 11. Cortaillod, Neuchâtel, Suisse (Von Burg 2002) - 12. Lastruccia, Sesto Fiorentino, Italie (Sarti et Martini 2001) - 13. Rubiera, Reggio Emilia, Italie (Bermond Montanari et al. 1982) - 14. Derrière-le-Château, Géovreissiat, Ain, France (Hénon et Vérot-Bourrely 1998) - 15. Les Calades, Orgon, Bouches-du-Rhône, France (Barge-Mahieu 1992a) - 16. Maupas, Vaunage, Gard, France (Roger 1992) - 17. Bois Sacré, Saint-Côme-et-Maruéjols, Gard, France (Roudil et al. 1974) - 18. Mourral, Trèbes, Aude, France (Vaquer 1998) - 19. San Martin de la Vega, Madrid, Espagne (Blasco et al. 1994) - 20. El Ventorro, Madrid, Espagne (Garrido-Pena 2000) - 21. Los Millares, Almeira, Espagne (Strahm 1995) - 22. Monte da Tumba, Alentejo, Portugal (Tavares da Silva et al. 1995) - 23. Zambujal, Estremadura, Portugal (Cardoso 2001). 40 Le Campaniforme... un phénomène... une énigme Dans la région du nord-est de la péninsule Ibérique, les habitats se localisent sur les hauts plateaux, sur les collines et en plaine. Des sites stratifiés ont été observés en grottes ou en abris, tels celui de Cova de Matadepera (Barcelone) ou de Balam del Duc (Rojalons) (Martin Colliga 1998). Dans la région de Valence, les habitats peuvent être regroupés en trois ensembles : les sites de vallée avec principalement des fosses-silos, les sites de hauteur, et les sites de plaine en construction en pierres (Marti Olivier et Juan-Cabanilles 2003). Les habitats peuvent être constitués de maisons ovales creusées dans le sol comme dans le site d’El Ventorro dans la région de Madrid. Elles peuvent également être rondes, en adobes, comme dans le site du Cerro de la Virgen (Los Millares) et des foyers ont également été mis en évidence à Picacès (Huesca). En Andalousie, les habitats campaniformes sont souvent des occupations successives, tels les sites de Los Millares et de Cerro de la Virgen (Martin Socas et al. 1998). Au Portugal, les sites se situent, en général, sur des collines ou plates-formes en éperons, souvent en bordure de vallée. Il s’agit en majorité d’une réoccupation de sites fortifiés présentant des conditions naturelles ou artificielles de défense, régulièrement agrandis lors de l’occupation campaniforme comme à Zambujal (Sangmeister et Schubart 1981), Monte da Tumba (Tavares Da Silva et Soares 1987), Vila Nova de Sao Pedro ou encore Leceia (Cardoso 1989, 2004). 1.2.4. La chronologie Le Campaniforme se développe pendant le 3e millénaire avant notre ère à l’articulation du Néolithique et du Bronze ancien. Plus de cent quatre-vingts datations radiocarbone sont accessibles permettant de dresser un aperçu du phénomène (Müller et van Willigen 1998). Ces dates ne résoudront pas l’énigme de l’origine du Campaniforme. En effet, outre les problèmes liés aux datations elles-mêmes - que nous ne débattrons pas ici -, celles-ci ne sont pas suffisamment nombreuses et sont inégalement réparties sur l’ensemble du territoire. Quoi qu’il en soit, les datations fournissent des éléments permettant de préciser quelque peu le contexte d’émergence et le développement du Campaniforme. En effet, l’exploitation des séries de dates radiocarbone permet de supposer que l’apparition du Campaniforme n’est pas simultanée en Europe et que ce phénomène se superpose à d’autres cultures, au moins matérielles. Ceci est prouvé non seulement par les dates, mais aussi par le mobilier. Il existe en effet dans plusieurs régions d’Europe des formes céramiques du Néolithique final à décor campaniforme et des formes céramiques campaniformes à décor Néolithique final (Bailly et Salanova 1999, p. 223). En Europe occidentale, les dates les plus anciennes se placent en 2900 et 2700 av. J.-C. Elles se situent en Europe méridionale (Portugal, Espagne, sud de la France). Plus on se dirige vers le nord et l’est, plus les dates sont récentes. En Suisse, en Italie et dans le nord de la France les premières datations ne se rencontrent pas avant 2500 av. J.-C. (Bailly et Salanova 1999). En Europe orientale, l’unique région qui dispose d’un nombre suffisant de datations est l’Elbe-Saale moyenne en Allemagne. Les datations les plus anciennes se situent dans la seconde moitié du 3e millénaire. Ailleurs, les rares datations permettent tout de même de confirmer le début du Campaniforme dès 2500 av. J.-C. L’analyse de l’Elbe-Saale permet également de percevoir une contemporanéité du Campaniforme avec les cultures Cordé et Unetice (Müller et van Willigen 1998). En Europe septentrionale, les datations semblent se situer dans la seconde moitié du 3e millénaire, voire plus tardives encore (Vander Linden 2006). Le Campaniforme prend fin aux alentours de 1800 av. J.-C. (Müller et van Willigen 1998). Les datations radiocarbone actuellement disponibles mettent en évidence un gradient sud-ouest - nord-est dans la mise en place du Campaniforme (Guilaine 1998c). Certaines datations ne s’intègrent pas à cette image car elles sont trop anciennes. C’est le cas pour une partie des dates radiocarbone des Pays-Bas, de la Hongrie et de l’Italie centrale. Pour M. Vander Linden (2006) ces différentes dates sont, respectivement selon les régions mentionnées ci-dessus, peu fiables, car sans contexte archéologique - donc inutilisables - et finalement imprécises. 41 Chapitre 1 1.3. Le Campaniforme hier Les recherches à la fin du 19e - début du 20e siècle concernent principalement la mise en place de la définition du complexe Campaniforme. Plusieurs chercheurs, comme E. Cartailhac (1886), P. Reinecke (1900), P. Bosch-Gimpera (1926), mettent en évidence les ressemblances et l’apparente homogénéité des vestiges retrouvés, notamment ceux de la céramique décorée. • Les explications diffusionnistes par la migration de populations Au départ, un consensus est partagé par l’ensemble des chercheurs : le Campaniforme est un phénomène migratoire. En revanche, la question de son origine et les voies de diffusion empruntées font l’objet de discussions plus ou moins animées. Pour certains chercheurs, comme O. Montelius (1900) ou J. Déchelette (1908), la source du Campaniforme est orientale, depuis l’Egypte avec une double voie de pénétration sur le continent par l’Italie et la Hongrie. L’Europe centrale est également proposée comme foyer potentiel d’apparition du Campaniforme. Pour d’autres, comme H. Schmidt (1913), P. Bosch-Gimpera (1926) ou encore A. Del Castillo (1928), l’origine ibérique du Campaniforme, en relation avec le nombre élevé de découvertes, ne fait aucun doute. G. Childe (1925), quant à lui, met en relation le déplacement d’individus avec une crise climatique ayant lieu à la fin du Néolithique, phénomène migratoire engageant des populations venant des steppes. J. Palliardi (1919) émet très tôt l’idée d’une origine double impliquant la péninsule Ibérique et l’Europe centrale dans les modalités de mise en place du Campaniforme. Dans le même esprit, E. Sangmeister (1963) propose, des années plus tard, la théorie du flux et du reflux (Rückstrom Theorie). Selon lui, l’origine du Campaniforme se place au Portugal. Une première migration a lieu par diffusion le long du littoral atlantique en direction septentrionale. Dans le nord et le centre de l’Europe, plusieurs phases d’acculturation se succèdent par confrontation avec la culture Cordé. Cette union donne naissance à un nouveau style qui se diffuse en laissant apparaître des styles régionaux. Finalement, une phase de reflux repart en direction méridionale en diffusant des éléments septentrionaux avec elle. A cette époque, on connaît avant tout les sépultures liées à ce complexe. Le matériel d’accompagnement laisse supposer que les individus rattachés au Campaniforme pourraient représenter des guerriers. On émet également l’hypothèse qu’il puisse s’agir de marchands, d’artisans ou tout simplement de nomades (Strahm 1998). De plus, les études concernant l’anthropologie physique corroborent les hypothèses d’un nouveau groupe d’individus possédant une morphologie bien distincte des populations locales (c.f. chapitre 1.5.). On retient parmi moult origines deux foyers probables : l’Europe centrale et la péninsule Ibérique, voire une situation mixte. On doit attendre les recherches conduites par J.N. Lanting et J.D. Van der Waals (1976) sur la typologie des céramiques décorées en relation avec les datations C14 aux Pays-Bas pour stabiliser momentanément la recherche. Pour ces auteurs, ce complexe est affilié au Cordé local - le groupe des sépultures individuelles - , puisque, dans cette région et dans le bas Bassin du Rhin, le Campaniforme le remplace de façon continue. Pendant longtemps, sur la base de leurs travaux, la quête de l’origine du Campaniforme semble enfin résolue et atténue les hypothèses diffusionnistes concernant la migration massive de populations comme unique explication du phénomène Campaniforme. • Les explications fonctionnelles par l’échange de biens socialement valorisés Dans les années soixante-dix, sous l’impulsion de la New Archaeology qui ne considère plus la culture matérielle comme étant un marqueur spécifique de groupes humains, les recherches sur le Campaniforme se réorientent. Ainsi, pour certains chercheurs, comme D.L. Clarke (1976) ou S.J. Shennan (1976), certains éléments du matériel associé au Campaniforme seraient en réalité des biens socialement valorisés - ou de prestige. Ils n’attesteraient pas de l’existence d’un groupe culturel particulier, mais se diffuseraient par des échanges entre élites au sein des cultures existantes. Pour D.L. Clarke (1976), la céramique décorée est un bien de luxe qui se transmet entre personnes de pouvoir. S.J. Shennan (1976), quant à lui, considère que 42 Le Campaniforme... un phénomène... une énigme l’ensemble de ces objets qu’il nomme package représente un assortiment de biens de prestige circulant au sein de communautés diverses. Pour R.J. Harrison, les raisons de l’apparition si brutale des gobelets, de leur rapide expansion et de leur succès général résident sûrement dans l’aggravation des différences de statut social à l’intérieur des sociétés de l’âge du cuivre. C’est l’accentuation de la stratification sociale et non pas les déplacements de marchands ou de nomades, qui fit de la poterie et des bijoux campaniformes des symboles enviés de richesse, et qui conduisit à leur adoption dans toute l’Europe à peu près à la même époque, vers 21002200. L’ « expansion » campaniforme a donc probablement été le résultat d’une compétition entre élites voisines rivalisant pour suivre les dernières modes en matière de richesse, dans le but d’exercer un contrôle supplémentaire sur les ressources alors disponibles (Harrison 1986, p. 141). • Le Campaniforme : un phénomène composite Dès 1979, A. Gallay développe un modèle dans lequel le Campaniforme est un phénomène composite. Selon cet auteur, il existe à la fois des mouvements complexes de populations et des phénomènes sociaux et commerciaux. Il propose six réseaux de relations regroupés en deux phases chronologiques (Gallay 1979, 2001). La première période correspond à la première phase d’expansion, depuis les Pays-Bas, des gobelets de type AOC dont les contacts et les échanges sont limités dans l’espace, soit en Grande Bretagne, en Bretagne et dans le Midi de la France (réseau 1), et à la diffusion des gobelets de type maritime sur la façade atlantique de l’Europe (réseau 2). La seconde phase (réseaux 3, 4, 5, 6) correspond à une différenciation régionale qui témoigne d’une rupture des réseaux d’échanges à grande distance présents lors de la première phase. Les deux premiers réseaux concernent la diffusion d’une mode, alors que les quatre derniers constituent, selon A. Gallay (1997-1998), autant de groupes culturels qui pourraient avoir chacun une origine différente. Cette situation n’exclut pas des contacts entre les groupes, ce qui expliquerait l’homogénéité des gobelets campaniformes. • Les explications idéologiques par l’expression emblématique du set Dans les années quatre-vingt dix, dans le cadre d’un séminaire3 sur le Campaniforme, C. Strahm et ses collaborateurs (1995) mettent en place un modèle qu’ils nomment Crémade. Le travail consiste, dans un premier temps, à l’élaboration d’un catalogue détaillé du matériel associé à cette manifestation. Ils individualisent différents groupes de vestiges (du matériel typiquement campaniforme à celui dérivé des substrats locaux) qui permettent, dans un second temps, d’inférer des postulats menant à un modèle explicatif de mise en place du Campaniforme. D’après leur classification, et le modèle Crémade qui en dérive, le Campaniforme est considéré comme l'expression d'une idéologie. Celle-ci résulterait de la volonté de certains individus d’exprimer leur indépendance et de se démarquer des autres (Benz et al. 1998). L’origine du phénomène se situerait en Europe centrale au sein des communautés locales, notamment dans celles du Cordé, et le Campaniforme représenterait une sphère d’échanges et de communications que l’on peut mettre en évidence à l’aide du set, nouvelle appellation du package défini par S.J. Shennan (1976). Ils s’appuient principalement sur le contexte de découvertes des éléments du set qui est presque exclusivement funéraire. Ainsi, pendant des décennies, les explications concernant l’apparition du Campaniforme varient entre mouvements de populations, circulations des biens ou des idées. Les recherches se basent avant tout sur le caractère paneuropéen des vestiges. De plus, elles ont souvent délaissé la variabilité certaine d’éléments moins exceptionnels de la culture matérielle au sein de leurs interprétations. 3 Le séminaire sur le Campaniforme dirigé par C. Strahm a été réalisé au sein de l’Institut für Ur- und Frühgeschichte de l’Université de Alberts-Ludwig à Freiburg-im-Breisgau (Allemagne). 43 Chapitre 1 1.4. Le Campaniforme aujourd’hui Les études touchant le Campaniforme ont ainsi souvent eu un caractère paneuropéen où l’accent était principalement mis sur les éléments originaux de la culture matérielle. Aujourd’hui, ce sont les différences qui sont recherchées au sein d’aires géographiques étendues par l’analyse d’instruments spécifiques. L’analyse des céramiques décorées à l’échelle européenne montre qu’il existe un gradient sud-ouest nord-est dans leur répartition (Salanova 2000, 2002). Pour L. Salanova, le standard est la céramique qui pose le problème campaniforme. Ils sont dispersés dans une grande partie de l’Europe, sans que des échanges de vases à longue distances ne soient clairement attestés (Salanova 2000, p. 193). En revanche, les céramiques décorées régionales témoignent seulement de la transformation sur place d’un style exogène (Salanova 2000, p. 193). Ainsi, pour cette auteure, le Campaniforme ne comporte que le standard. Les études de la pétrographie des poteries sont intéressantes à plus d’un titre. Elles sont unanimes et indiquent, indifféremment des régions étudiées, que les céramiques sont largement produites sur place et un nombre restreint de poteries font réellement l’objet d’importations (Rehman et al. 1992 pour la République tchèque et la Hongrie, Millán et Arribas 1994 et Clop 2002 pour l’Espagne, Convertini 1996 pour la France et la Suisse...). De plus, les composantes argileuses des céramiques montrent un gradient sud - nord dans la proportion relative de la chamotte comme dégraissant (Convertini 1996). Une étude concernant la mise en place du Campaniforme dans le sud-est de la France par l’analyse de la céramique décorée permet de proposer un schéma de développement de ce complexe dans cette région (Lemercier 2002, 2004). La première étape serait constituée par l’implantation de groupes campaniformes clairement identifiés par leur culture matérielle au sein des cultures locales du Néolithique final. Ces groupes diffuseraient les vases du standard autour de leurs implantations et jusque dans des régions où ils ne sont pas directement implantés. Elle serait suivie du développement d’une réelle culture matérielle campaniforme régionale par acculturation des groupes locaux, intégrant un ensemble culturel vaste mais développant leur faciès local (Lemercier 2002, p. 228). L’étude de l’industrie lithique de cette même région permet également de mettre en évidence un phasage bipartite avec un stade ancien très hermétique et une phase récente présentant une industrie lithique plus hétérogène, où il est possible de noter les influences avec les traditions locales du Néolithique final (Furestier 2005). Pour cet auteur, ces résultats peuvent être étendus à l’ensemble de l’Europe même si ce phasage bipartite est moins évident (Furestier 2005). L’analyse des céramiques communes de ce groupe culturel permet aujourd'hui de définir trois domaines (oriental, septentrional et méridional) pour lesquels les substrats néolithiques précampaniformes jouent des rôles distincts selon les régions (Besse 2003). En effet, des liens étroits existent entre le Cordé et le Campaniforme en Europe centrale. Il n’en est pas de même dans le Bassin méditerranéen, où les céramiques montrent une rupture plus marquée par rapport aux divers groupes culturels précampaniformes. Ainsi pour l’auteure, le Campaniforme est [...] caractérisé par des réseaux complexes de diffusion et des mécanismes de transition à partir du substrat très différents selon les aires retenues où il est possible de percevoir des axes d’influences privilégiés (Besse 2001, p. 76). Les traits culturels opposant les domaines oriental et méridional sont également soutenus par d’autres composantes culturelles. L’étude de l’industrie lithique montre que les pointes de flèches triangulaires trapues sont à rattacher au complexe oriental ; en revanche, les armatures de flèche fusiformes ou encore pédonculées et à ailerons sont à attribuer à l’aire méridionale du phénomène (Bailly 2002). On le voit bien, aborder le Campaniforme sur l’ensemble du territoire qu’il occupe, n’est pas une tâche aisée! Cependant, ces différentes études permettent de constater qu’on ne peut plus aujourd’hui envisager un seul schéma comme unique explication du phénomène Campaniforme. Les modalités de l’émergence de ce complexe possèdent de multiples facettes des plus difficiles à maîtriser comme un tout. 44 Le Campaniforme... un phénomène... une énigme 1.5. Et quand l’individu s’exprime... associé à la culture Campaniforme Et quand l’Homme associé à la culture Campaniforme se manifeste, que dit-il? Les premières études le concernant ont débuté à la fin du 19e - début 20e siècle. Elles ont abordé sa morphologie crânienne. On peut citer, entre autres, les travaux de D. Wilson (1851) sur les mesures crâniennes des Campaniformes de Grande Bretagne ou encore les études de A. Schliz (1910) et de K. Gerhardt (1953, 1976) touchant essentiellement les données provenant de l’Allemagne. Ces études anthropométriques sont des classifications purement typologiques et descriptives. Elles mettent en évidence une opposition marquée entre la morphologie particulière des Campaniformes et celle des populations de la fin du Néolithique. Ces dernières ont une morphologie franchement dolichocéphale (crâne allongé) ; en revanche, les Campaniformes sont majoritairement brachycéphales, plannocipitaux et eurymorphes, - en d’autres termes, présentant des crânes arrondis avec un aplatissement de l’occipital et une face large. Dans les années soixante-dix, l’informatique engendre la réalisation d’analyses multivariées sur les données métriques, élément non négligeable permettant le dépassement d’études exclusivement descriptives. R. Menk (1979, 1981) propose de ce fait une synthèse à l’échelle européenne du complexe Campaniforme. Il dispose de plus de 1200 individus dont 350 campaniformes, et les sujets restant se partagent entre le substrat précampaniforme et le Bronze ancien. L’échantillon étudié rassemble 37 ensembles provenant d’Allemagne, de Suisse, d’Italie, d’Espagne, du Portugal, de Tchéquie, de Pologne et de Grande-Bretagne. Il utilise 16 mesures et 6 indices crâniens avec lesquels il effectue des analyses multivariées. Il recherche, dans un premier temps, la supposée uniformité campaniforme. Ses résultats plaident en faveur d’une humanité campaniforme qui se rencontre, à quelques exceptions près, partout où l’occupation campaniforme est attestée. Par humanité, il se réfère au type morphologique caractéristique des Campaniformes : la brachycéphalie planoccipitale eurymorphe. Il définit une zone nucléaire, où la morphologie est homogène, composée de l’Allemagne centrale, la Moravie, la Bohème et la Pologne ; en périphérie, par contre, le type campaniforme devient minoritaire et diminue en fonction de l’éloignement de la zone centrale. Il s’intéresse, dans un deuxième temps, à l’origine de cette humanité campaniforme. Celle-ci n’est pas rattachable, selon lui, à un substrat local rencontré dans l’aire de répartition examinée. Son origine est probablement à chercher en Europe du sud-est en liaison avec l’expansion de la civilisation des Kourganes. Ensuite, il s’attache à la participation du Campaniforme dans la formation des civilisations du Bronze ancien. Mais, selon ses résultats, l’humanité campaniforme s'estompe dans la presque totalité des régions touchées par l’occupation campaniforme pendant le Bronze ancien. Finalement, il réalise des analyses individuelles par région afin de cerner plus précisément les modalités de mise en place de complexe. Nous verrons en détail ses résultats selon les régions que nous avons sélectionnées dans notre étude dans le chapitre 2. Les résultats de l’anthropométrie s’intègrent parfaitement aux interprétations issues des données archéologiques de l’époque. Ainsi, pour l’archéologie et l’anthropologie, l’apparition du Campaniforme est liée à la circulation de personnes. Plus tard, les débats s’animent et l’idée que le Campaniforme soit la conséquence de mouvements de populations est très vite abandonnée au profit de la diffusion des composantes culturelles. Les résultats de l’anthropométrie sont remis en cause et un nouveau consensus semble rassembler la plupart des chercheurs: la morphologie subit d’importants effets environnementaux et devient de ce fait un indicateur inadapté de distances biologiques entre les populations. Les chercheurs s’appuient notamment sur les travaux de F. Boas (1912). En effet, cet auteur étudie les variations de l’indice céphalique, mesure sur le vivant correspondant à l’indice crânien, sur un échantillon de plus de 13'000 individus constitué d’immigrés et de leurs descendants nés aux Etats-Unis. Sa recherche permet d’affirmer que la morphologie crânienne n’est pas uniquement héritée et que l’environnement joue un rôle - quoique modéré - dans son déterminisme. Dès lors, les interprétations iront au-delà des 45 Chapitre 1 conclusions émises par l’auteur lui-même jusqu’à imaginer une plasticité totale de la forme du crâne (Holloway 2002). En 1983, E. Kobyliansky (1983) effectue une recherche similaire en étudiant quatre groupes d’immigrants, dont un échantillon constitué d’individus apparentés sur trois générations (une lignée migrante et deux descendances). Ses résultats vont dans le sens de ceux de F. Boas. Des modifications significatives de l’indice céphalique peuvent être constatées à l’issue d’une ou deux générations seulement. N. Brodie (1994), quant à lui, réalise une critique des outils méthodologiques en archéologie et en anthropologie aboutissant aux interprétations touchant la mise en place du Campaniforme en Grande Bretagne. Il consacre une grande partie de son étude à l’anthropométrie et la signification des changements de la morphologie crânienne. Il met en relation les variations de l’indice crânien de ces populations avec des variations climatiques ayant lieu à la fin du Néolithique. Ainsi, pour cet auteur, un climat plus sec avec une élévation de la température de 1 à 2°C, entraîne des modifications de l’alimentation, elles-mêmes corrélées au degré de brachycéphalie (Brodie 1994). Toutes ces recherches mettent en avant l’influence de l’environnement sur la morphologie crânienne. Malgré une abondante littérature sur le sujet (Susanne 1977, Devor et al. 1986...), il aura fallu attendre des années pour relancer véritablement le débat sur la présumée plasticité du crâne. En effet, ces dernières années, deux groupes de chercheurs - C.S. Sparks et R.L. Jantz (2002, 2003) et C.C. Gravlee, H.R. Bernard et W.R. Leonard (2003a/b) - ont repris les données de F. Boas à l’aide de nouveaux outils statistiques. Ils ont a priori travaillé sur les mêmes données en employant différentes analyses mais leurs résultats sont en pleine contradiction. Pour C.S. Sparks et R.L. Jantz (2002, 2003), la composante génétique de la forme du crâne est très forte. A l’opposé, les résultats de C.C. Gravlee et collaborateurs (2003a/b) vont dans le sens des interprétations de F. Boas en confirmant une certaine plasticité de la morphologie crânienne. On le voit bien, relancer le débat ne signifie pas trouver la solution. La question n’est donc toujours pas résolue. A l’évidence, there is more to know about genetic and environmental influences that might vary in different populations [...] Legions of us are dying to know the answers (Holloway 2002, p. 14623). Pour R.L. Holloway (2004), l’indice crânien semble avoir une forte composante génétique, tout en étant légèrement influencé par l’environnement (le régime alimentaire, la santé, peut-être le climat). Ces influences externes ne parviendront jamais à transformer des têtes longues en têtes larges, pas plus que l’inverse. [...] Il est donc temps de tordre le coup à l’idée que la forme de la tête puisse être entièrement modelée par l’environnement (Holloway 2004, p. 59). Au-delà de cela, d’autres éléments viennent se superposer dans l’énigme posée par le Campaniforme. Le problème de la morphologie crânienne ne se résume pas seulement à la recherche de l’apport respectif des gènes et de l’environnement dans le déterminisme de sa forme. Il faut également évoquer l’adéquation réalisée pendant des décennies entre la brachycéphalie et le Campaniforme. Qui n’a jamais entendu si cet individu est brachycéphale, alors il s’agit d’un Campaniforme ainsi que sa réciproque ? Des réévaluations d’échantillons permettent de préciser la relation entre le Campaniforme et la brachycéphalie. Un bilan des données crâniennes des populations du Néolithique de la Suisse occidentale permet de nuancer le schéma généralement adopté des populations du Néolithique moyen à morphologie dolichocéphale - crâne allongé -, de celles du Néolithique récent à morphologie mésocéphale - crâne de forme intermédiaire - et de celles campaniformes à morphologie franchement brachycéphale - crâne arrondi - (Desideri à paraître). On trouve déjà en abondance la forme intermédiaire, la mésocrânie au Néolithique moyen et la majorité des individus sont brachycrânes au Néolithique récent. La relation ici entre la brachycéphalisation et l’émergence du Campaniforme ne se justifie pas. On peut également citer les nouvelles déterminations d’individus brachycéphales de la Basse vallée du Rhône (France) attribués initialement au Campaniforme et se révélant appartenir au Bronze ancien (Tchérémissinoff 2006). Les polémiques autour de la morphologie crânienne n’ont pas entraîné son abandon au sein de la communauté anthropologique (Černý 1996, Eades 1996...). Le caractère brachycéphale des individus campaniformes est souvent mis en avant, mais lui donner un sens reste de nos jours encore un sujet délicat. Les chercheurs ont des difficultés à légitimer la morphologie particulière des individus 46 Le Campaniforme... un phénomène... une énigme campaniformes par l’apport de nouvelles populations, ils préfèreront parler d’évolution interne des sociétés (notamment Černý 1996 pour la Bohême). De nouvelles approches, moins discutées que la métrique, se développent au sein des études anthropologiques touchant la mise en place du Campaniforme (Czarnetzki 1984, Eades 1996, Desideri 2001, Desideri et Eades 2002, 2004). Les analyses concernant les variations morphologiques non métriques du crâne et de la dentition prennent leur envol. Ces caractéristiques morphologiques sont considérées comme de bons indicateurs des relations biologiques entre les populations (c.f. chapitre 4). Les résultats mettent en évidence, selon les régions étudiées, des différences importantes des ensembles liés à ce complexe qui pourraient être mises en parallèle avec l’arrivée de nouvelles populations, ou alors à un accès modifié au sein des aires funéraires (Desideri et Eades 2002, 2004 pour la Suisse), voire des comportements différents selon le sexe des individus campaniformes (Czarnetzki 1984 pour la Bohême). En parallèle, le Campaniforme bénéficie de quelques études très intéressantes concernant la géochimie isotopique (Price et al. 1998, 2004, Chiaradia et al. 2003, Fitzpatrick 2003, Evans et al. 2006...). Les signatures isotopiques osseuses s’avèrent un bon outil dans la restitution de l’histoire des peuplements à condition de pouvoir exclure tout problème de diagenèse. Certains auteurs affirment que postdepositional contamination of bone and tooth is normally not a significant problem in strontium isotope studies (Price et al. 2004, p. 15). Ces taux peuvent servir d’indicateurs des aires géologiques dans lesquelles un individu a vécu, car les éléments chimiques sont assimilés dans le squelette pendant sa croissance et son remodelage. L’os subit un remplacement continuel de ses fractions inorganiques, c’est pourquoi les mesures de la composition isotopique osseuse reflètent les dernières années de vie d’un individu. Ces éléments vont également s’incorporer dans la fraction minérale des dents durant toute la formation de la dentition. Puis, après l’éruption des dents, contrairement à l’os, la composition chimique de l’émail ne subit que très peu de modifications. Elle reflète donc les conditions extérieures au moment de sa formation. Ainsi, les taux de compositions isotopiques entre les os et les dents permet de refléter les changements dans l’histoire de résidence d’un individu. Ces études appliquées au Campaniforme enregistrent une certaine mobilité des individus liés à ce complexe (Price 1998, 2004, Chiaradia et al. 2003), voire même une origine allochtone de ceux-ci (Fitzpatrick 2003, Evans et al. 2006). Pour l’Europe centrale, les analyses chimiques de 81 individus montrent des mouvements de populations pendant le Campaniforme sur l’ensemble de l’aire étudiée (Price et al. 2004). En effet, selon la méthode employée, une partie des individus étudiés ne serait pas d’origine locale (60% ou 17% selon l’analyse). Cette mobilité ne semblerait influencée ni par l’âge, ni par la chronologie interne du Campaniforme ; que l’on soit dans une phase ancienne ou récente du phénomène n’influe pas sur les résultats. Par contre, les femmes sembleraient tout de même être slightly more mobile (Price et al. 2004). Dans une perspective plus éloignée de la dynamique du peuplement, des études sur la biomécanique ont vu le jour touchant la caractérisation de la vie quotidienne des individus. La Bohême est une région qui regorge de structures funéraires mais se distingue par un nombre restreint de découvertes domestiques attribuées à la fin du Néolithique. Certains archéologues, comme M. Buchvaldek (1985) ou S. Vencl (1994), interprètent cette déficience par une économie de subsistance pastorale et nomade de ces populations. L’analyse de la distribution et de la forme des sections transverses des os longs, que l’on regroupe sous le terme de propriétés géométriques des sections transverses, peut fournir des clés de compréhension pour valider ou non ces hypothèses. Dans ce cas précis, la biomécanique ne constate aucune mobilité significative des individus de la fin du Néolithique de Bohême qui pourrait être mise en parallèle avec l’existence de communautés pratiquant le nomadisme pastoral (Sladek et al. 2006a/b). 47 Chapitre 1 1.6. Un programme interdisciplinaire... un objectif commun La caractérisation du Campaniforme se révèle un exercice des plus laborieux. Aborder dans son entité géographique totale ce complexe semble néanmoins indispensable si l’on souhaite affiner et préciser les connaissances que l’on peut en avoir. Sur la base d’une première recherche concernant l’étude de la céramique commune (Besse 2001, 2003, 2004a/b) qui a permis de révéler, comme nous l’avons vu précédemment, des comportements différents sur l’ensemble du territoire occupé par la culture Campaniforme, un programme interdisciplinaire dirigé par la professeure M. Besse et financé par le Fonds National de la Recherche Scientifique (FNS requêtes 100012-100599 et PP001-102710) a vu le jour. Ce programme vise à une meilleure compréhension du Campaniforme par l’étude d’une aire géographique large. L’objectif principal étant de saisir les mécanismes impliqués dans le passage du Néolithique final précampaniforme au Campaniforme par l’étude de quatre volets complémentaires. • La céramique commune et les datations radiocarbone La première partie du programme concerne l’étude de la céramique commune de plus de neuf cents sites, localisés en Allemagne, en Autriche, en Belgique, en Espagne, en France, en Hongrie, en Italie, aux PaysBas, en Pologne, au Portugal, en République tchèque, en Slovaquie et en Suisse4. La céramique commune étudiée peut être définie par quatre-vingt-trois types, dont vingt-six constituent les types les plus représentés. Parmi les types principaux de céramiques, les plus pertinents sont sélectionnés afin de déterminer leur contexte d’apparition, soit au Campaniforme, soit dans les groupes précampaniformes, sur la base des datations radiocarbone associées. Afin de pouvoir y répondre, ce volet aborde différents points. Dans un premier temps, il s’agit de compléter les données déjà acquises pour les pays mentionnés ci-dessus en poursuivant l’inventaire des sites campaniformes ayant livré de la céramique commune. Dans un deuxième temps, il s’agit d’étudier les principaux types de céramiques communes présents dans plus de 900 sites et de les mettre en relation avec les datations radiocarbone, en contrôlant l’association entre l’échantillon daté et le type céramique retenu. Il est possible par la suite de situer temporellement la présence de chaque type céramique au sein du Campaniforme. En dernier lieu, il faut définir si les types principaux de céramiques communes sont présents dans les groupes précampaniformes et, si tel est le cas, en déterminer l’origine et la position spatiotemporelle pour chacun d’entre eux. • L’occupation du territoire Cette étude aborde la diversité des sites campaniformes sur l’ensemble de l’Europe, tant au niveau des structures domestiques que des funéraires 5. Une première partie de ce volet synthétisant la diversité des structures domestiques et funéraires associées au Campaniforme vous a été présentée auparavant par les figures 6 et 7. Il est une évidence : des sépultures individuelles au monument mégalithique, de la délocalisation de l’habitat campaniforme à la proximité ou à la réoccupation de celui du Néolithique final, les faits archéologiques sont peu comparables d’une région à l’autre de l’Europe. La seconde partie de ce volet procède à l’évaluation des composantes culturelles identitaires émanant d’un choix de celles répondant à des contraintes (notamment techniques et mésologiques). Et l’habitat, dans son acceptation la plus large, en fait un excellent document. L’objectif étant de mettre en évidence la part du culturel de celle des contraintes par l’exploitation de certains constats portant sur la localisation des sites campaniformes. Les habitats campaniformes sont construits soit sur un territoire « vierge », soit à proximité ou sur des habitats du Néolithique final. La question est dès lors de savoir s’il s’agit de la manifestation d’une volonté délibérée de se démarquer des populations du Néolithique final, et le cas échéant, d’en saisir les raisons. 4 5 48 Ce volet est réalisé par la professeure M. Besse et par M. Piguet. Ce volet est réalisé par la professeure M. Besse. Le Campaniforme... un phénomène... une énigme • La métallurgie du cuivre Cette étude vise à mieux comprendre les modalités d’appropriation d’une nouvelle matière première, le cuivre, et des technologies associées sur le domaine alpin pendant le 3e millénaire avant notre ère6. Parallèlement, le Valais a été choisi pour mener une étude sur l’exploitation du minerai de cuivre du Néolithique final au Bronze ancien. Le Campaniforme a donc une place de choix dans cette recherche. Si les connaissances sur la métallurgie de l’Arc alpin, pris dans sa globalité, sont acquises sur une base bibliographique, la détermination d’une exploitation (ou non) du minerai de cuivre valaisan se base sur du matériel archéologique du Valais central et du Plateau suisse. La méthode repose sur la comparaison des signatures isotopiques du plomb et chimiques d’un corpus d’objets en cuivre et en bronze avec les signatures de minerais de cuivre valaisans. • L’anthropologie biologique Le dernier volet correspond à notre propre recherche et fait l’objet de ce travail afin d’acquérir les données de l’anthropologie biologique. La mise en parallèle et en commun des observations fournies par ces différentes approches devrait permettre d’avoir une meilleure connaissance de l’émergence du complexe Campaniforme, des faits archéologiques à la dynamique du peuplement. 1.7. Les objectifs de cette recherche Explanations for the Beaker phenomenon advanced since the 1960s have tended to be mainly or entirely built around the artifactual data. Osteological evidence is rarely discussed on an equal footing... (Mays 1998, p. 97) Si l’individu a été délaissé des interprétations concernant la compréhension du Campaniforme, il reste néanmoins l’un des témoins privilégiés d’une culture que l’on a finalement du mal à définir dans son entièreté. Il peut sans aucun doute fournir des clés de compréhension des plus intéressantes. En effet, pour saisir si les transformations liées à l’apparition du Campaniforme peuvent être attribuées à l'arrivée de nouvelles populations, l’individu doit non seulement pouvoir s’exprimer mais également être analysé. La volonté de faire parler les individus qui sont à l’origine du Campaniforme n’est pas inédit. De l’étude locale à l’échelle du phénomène, de la morphologie du crâne à la composition chimique des ossements, les travaux ont souvent mis en évidence une certaine mobilité des individus campaniformes. Elle est définie comme partielle, totale ou encore différentielle. La morphologie crânienne - la brachycrânie à aplatissement occipital - caractéristique des individus campaniformes en est la raison. Pendant des décennies, cette morphologie a été considérée comme une preuve de mouvements de populations pendant le Campaniforme. Mais les variations perpétuelles des interprétations issues des composantes matérielles ont attribué à l’étude des individus - ainsi qu’à ses résultats - une position de dernier de la classe. Pourtant, dans cette quête inlassable d’éléments permettant de retracer les modalités de mise en place du Campaniforme... l’anthropologie physique reste un partenaire incontournable... (Vander Linden 2006, p. 163). Ainsi, pour appréhender à nouveau les modalités d’apparition du Campaniforme par l’intermédiaire des individus, il est indispensable de choisir des éléments sur le squelette humain susceptibles de fournir des éléments de réponse intéressant la dynamique de peuplement. Nous avons retenu la dent comme outil d’étude et, plus particulièrement, les variations anatomiques observées sur l’ensemble des dentitions permanente et déciduale. 6 Ce volet fait l’objet d’une thèse de doctorat intitulée La métallurgie du cuivre dans les Alpes occidentales au 3e millénaire avant J.-C. réalisée par F. Cattin (Cattin en cours). 49 Chapitre 1 Nous avons sélectionné plusieurs territoires à occupation campaniforme. Si les régions les plus représentatives et les plus importantes impliquées dans l’émergence de cette culture ont d’abord été retenues, le choix définitif s’est fondé sur la disponibilité et l’accessibilité des collections anthropologiques. Nous parlerons ici de domaines en référence aux résultats de l’étude de la céramique commune (Besse 2001, 2003, 2004a/b). Ainsi, cinq territoires sont étudiés : la Bohême et la Hongrie pour le domaine oriental, le sud de la France et le nord de l’Espagne pour le domaine méridional et la Suisse à l’articulation entre les deux domaines. L’objectif est de définir, pour chacune de ces régions, si l’émergence du Campaniforme coïncide ou non avec un renouvellement de la population. Nous aborderons des points précis, par l’analyse de la morphologie dentaire de ces populations, qui devraient permettre de restituer leur histoire à travers le temps. Dans un premier temps, nous souhaitons avoir une vision locale des relations entre les populations de la fin du Néolithique. Divers aspects seront ainsi traités : • Les relations entre les ensembles précampaniformes Il s’agit de déterminer le comportement des différents groupes locaux précédant le Campaniforme et leur interaction. • Les relations entre les ensembles du Campaniforme Nous nous intéressons ici aux rapports entretenus entre les différents ensembles locaux rattachés à la culture Campaniforme. • Le rôle du fonds précampaniforme local dans la mise en place du Campaniforme Sur la base des points précédents, nous tenterons de déterminer si l’on a affaire à une rupture de peuplement ou non. • Le rôle du fonds campaniforme dans la mise en place du Bronze ancien Il s’agit de comprendre si le Campaniforme est ou non responsable de la mise en place du Bronze ancien. Selon les spécificités des échantillons composant les différentes régions étudiées, nous pourrons répondre intégralement ou seulement partiellement à ces différents points. Dans un second temps, nous aborderons les connexions entre les populations sur une aire géographique étendue. Deux points seront approchés : • Les relations entre les groupes du Campaniforme sur l’ensemble du territoire étudié Nous nous attacherons aux groupes régionaux campaniformes et aux affinités qu’ils pourraient éventuellement entretenir entre eux. • La dynamique du peuplement Nous souhaitons saisir les interdépendances entre les différents groupes culturels. Par une vision spatiotemporelle large, nous aborderons l’histoire de ces populations et leur relation. Pour répondre à ces différentes questions, nous réaliserons, pour toutes les aires étudiées, une analyse individuelle de chaque région (Bohême, Hongrie, sud de la France, nord de l’Espagne et Suisse), puis nous effectuerons une analyse de chaque domaine (oriental, méridional et à l’articulation - la Suisse -). Nous poursuivrons ensuite les analyses par une étude comprenant uniquement les échantillons campaniformes de cinq régions traitées. Nous terminerons la phase analytique par une étude sexes séparés que sera réalisée sur les échantillons tchèques uniquement, puisque la Bohême est la seule région pour laquelle nous avons des déterminations sexuelles suffisantes. Finalement, il sera intéressant et utile de discuter des résultats apportés par la morphologie dentaire et de les confronter aux interprétations issues d’autres approches complémentaires - anthropologie biologique, archéologie, pratiques et rituels funéraires… Nous tenterons non seulement de dresser un bilan des modalités de mise en place du Campaniforme, mais également de suggérer un scénario d’évolution de la société pendant le 3e millénaire à l’échelle européenne. 50 Le Campaniforme... un phénomène... une énigme 1.8. L’essentiel en bref Ce premier chapitre a abordé l’essence même de ce travail : le Campaniforme. Nous nous sommes intéressée à ce qui fait le Campaniforme, à ce qu’on en a dit hier et ce qu’on en pense aujourd’hui. Nous avons également situé notre recherche au sein d’un programme interdisciplinaire abordant l’histoire du peuplement du 3e millénaire avant notre ère. Nous avons finalement énoncé les objectifs de notre recherche. Nous avons vu que : • Le Campaniforme se met en place à l’échelle européenne sur des substrats locaux très différents. De l’unité précampaniforme orientale à l’hétérogénéité occidentale, les faits sont peu comparables. • Le Campaniforme est caractérisé par un matériel composé d’une céramique décorée - dénominateur commun de cette culture -, d’une céramique commune variant selon les régions, d’un matériel divers constitué notamment de brassards d’archer, de poignards en cuivre, de pointes en cuivre dite de Palmela, de boutons en os perforés en V... et de très belles stèles anthropomorphes. • Si le mobilier fédère le Campaniforme, la diversité incontestable de ses structures funéraires et domestiques le brouille. Ainsi, de la sépulture individuelle, à la réutilisation ou à la pratique de tombes collectives, en passant par l’incinération, le rituel funéraire des Campaniformes nous révèle ici toute sa complexité. De même, la variabilité des structures domestiques, tant au niveau de leur construction sur poteaux ou en pierres sèches -, qu’à leur forme - circulaire, ovalaire, rectangulaire -, ou encore à leur localisation - à proximité ou non de leur substrat-, est certaine. • Le Campaniforme se développe pendant le 3e millénaire avant notre ère, entre 2900 et 1800 av. J.-C., et il est possible de mettre en évidence un gradient sud-ouest - nord-est dans son émergence. • Les premières explications concernant l’émergence du Campaniforme s’appuyaient principalement sur le caractère paneuropéen de certains vestiges et ont variés entre mouvements de population, circulations des biens ou encore des idées. • Aujourd’hui, les recherches s’intéressent aux divergences. Ce sont de vastes territoires qui sont analysés à l’aide d’instruments spécifiques. Les éléments moins exceptionnels de la culture matérielle comme la céramique commune ou encore l’industrie lithique s’expriment et mettent en évidence une certaine dichotomie du Campaniforme opposant l’est et l’ouest du phénomène. • Les études anthropologiques ne sont pas en reste, de l’étude ponctuelle à l’échelle européenne, de l’observation de la forme du crâne à la composition chimique des ossements. La mobilité, - partielle, totale ou différentielle - selon l’approche employée, est souvent associée à l’émergence du Campaniforme. La morphologie crânienne particulière des individus campaniformes en est à l’origine - la brachycrânie à aplatissement occipital. Cette morphologie a longtemps été considérée comme un indice certain de mouvements de populations à l’origine du Campaniforme. Aujourd’hui, d’autres approches, moins sujettes à discussion, comme la géochimie isotopique des os ou encore les traits non métriques sont privilégiées. • Notre recherche s’intègre au sein d’un programme interdisciplinaire qui a pour objectif la compréhension des mécanismes impliqués dans l’émergence du Campaniforme par l’étude de quatre volets complémentaires : la typologie et chronologie de la céramique commune, l’occupation du territoire, la métallurgie et l’anthropologie dentaire. Ces différents éléments permettent de définir les objectifs de notre recherche : avons-nous affaire à une rupture ou à une continuité de peuplement lors de l’apparition du Campaniforme ? Et pour répondre à cette question, cinq régions - la Bohême, la Hongrie, le sud de la France, le nord de l’Espagne et la Suisse - ont été sélectionnées représentant la plupart de la variabilité de l’expression du Campaniforme et totalisant plus de 2’000 individus, 530 traits et 255’000 observations. 51 Chapitre 2 Du groupe culturel à l’individu 2.1. Les régions sélectionnées Nous avons vu que le Campaniforme se développe sur l’ensemble de l’Europe de manière non uniforme. Nous avons ainsi sélectionné cinq régions qui, inévitablement, ne représentent pas l’ensemble de la variabilité de l’expression du Campaniforme, mais du moins, une large partie de celle-ci. Ces territoires concernent deux des trois domaines définis par l’étude de la céramique commune (Besse 2001). Deux régions appartiennent au domaine oriental. La première, la Bohême, est une région importante pour la compréhension du Campaniforme, non seulement par son nombre élevé de découvertes, mais également par le rôle qu’elle a probablement joué dans la mise en place de ce groupe culturel. Nous considérons pour cette région son substrat, en partie contemporain, le Cordé, et la période qui lui succède, la culture d’Unetice, le Bronze ancien. La seconde région du domaine oriental est la Hongrie. Le substrat sur lequel le Campaniforme se développe est la culture de Makó, qui ne sera pas inclus dans les analyses puisque nous n’avons pas eu la possibilité d’en observer des échantillons. Ainsi, nous abordons cette région avec des cultures qui sont soit contemporaines, soit successives au Campaniforme. L’aire méridionale est un territoire où les modalités de mise en place du Campaniforme seront, en considérant, d’une part, la complexité de l’expression de ce phénomène, d’autre part, la nature de ses vestiges, probablement plus difficiles à saisir. Deux régions concernent ce domaine. La première est le sud de la France pour laquelle nous avons réuni des ensembles appartenant au Néolithique final, au Campaniforme et au Bronze ancien. La seconde correspond au nord de l’Espagne, région pour laquelle nous étudions des échantillons appartenant au Néolithique final et au Chalcolithique avec ou sans Campaniforme. Le Bronze ancien, trop peu représenté, ne pourra pas être intégré dans les analyses. La dernière région sélectionnée est la Suisse qui se situe à l’articulation entre ces deux grands courants. Sa position intermédiaire se reflète également dans la diversité de ses gisements, même s’ils ne sont pas nombreux. La Suisse orientale s’oppose ainsi à celle occidentale, du moins au niveau du rituel funéraire, l’est suivant la tradition des inhumations individuelles et l’ouest celle des tombes collectives. Le territoire helvétique possède une séquence funéraire sans hiatus entre le Néolithique moyen et le Bronze ancien, de ce fait nous abordons cette région dans une perspective chronologique élargie par rapport aux autres régions étudiées. Figure 8 : Les régions étudiées dans ce travail. Le domaine septentrional, quant à lui, n’est pas intégré dans notre étude pour des raisons diverses et variées. D’une part, les vestiges osseux ne se retrouvent pas en suffisance, ce qui aurait fortement limité nos analyses. De l’autre, la réunion des ensembles étudiés ici a demandé un temps et une énergie considérables. Aborder le domaine septentrional avec sa complexité observée sur les données archéologiques est un sujet de thèse en soi. Par conséquent, ceci a restreint notre sélection pour concentrer nos efforts sur certaines régions seulement. Une présentation de tous les sites et de chaque individu7 intégrés dans notre travail se trouve respectivement en annexes 1 et 2. 7 Nous considérons ici aussi bien les sujets individualisés, les lots de dents en vrac que les dents isolées des sépultures collectives. 53 Chapitre 2 2.2. Le domaine oriental : la Bohême La Bohême est une région importante dans la compréhension du Campaniforme. Elle a été considérée à plusieurs reprises dans l’histoire des recherches comme un potentiel foyer d’apparition du Campaniforme, ou du moins d’un certain Campaniforme (Harisson 1974, Besse 2004b, Vander Linden 2004…). 2.2.1. Le Campaniforme en Bohême : son insertion culturelle, spatiale et temporelle En Bohême, l’Enéolithique est une période de développement indépendant plus proche du Bronze ancien que du Néolithique dans ses caractéristiques générales (Zapotocký 1994). Le Campaniforme apparaît au sein de sa phase finale (Late Eneolithic). Ce dernier est subdivisé en trois étapes - le Cordé, le Campaniforme et le proto-Unetice. Il ne s’agit en aucun cas de la succession de trois périodes distinctes puisqu’elles se superposent les unes aux autres. En effet, le Campaniforme coexiste tantôt avec le Cordé, tantôt avec le proto-Unetice. Le Cordé de Bohême est sans doute le groupe culturel le mieux connu de la fin du Néolithique oriental, cependant sa genèse reste toujours problématique (c.f. chapitre 1.1.). Dans cette région, la distribution du Campaniforme est parfois conditionnée par l’implantation du Cordé. En effet, ces deux ensembles s’excluent dans le nord-est de la Bohême (Vander Linden 2006). Ceci n’est pas le cas dans le nord-ouest et en Moravie puisque le Campaniforme dans ces régions supplée le Cordé. Le proto-Unetice marque, quant à lui, la fin de l’Enéolithique sans interruption avec le Bronze ancien (Zapotocký 1994). L’Enéolithique final débute aux alentours de 2900-2700 av. J.-C. et se termine - vers 2400 av. J.-C. - avec l'établissement du Bronze ancien (la culture d’Unetice) (Zapotocký 1994). 2.2.2. Le Campaniforme en Bohême : les données de l’archéologie Une chronologie en trois phases interne au Campaniforme a été établie sur la base des assemblages céramiques (Hájek 1966, Neustupný E. 1984). La durée de ces phases ne peut être déterminée précisément. La phase ancienne se compose uniquement de céramiques décorées, dont le décor est fait de fines bandes ornées en alternance avec des bandes vides. Les tombes de la phase ancienne sont souvent isolées ou situées dans de petites nécropoles. La phase moyenne regroupe des céramiques décorées dont le décor s’organise en larges bandes ainsi que de céramiques non décorées. La dernière phase comporte uniquement ce dernier type de céramique et est relativement mal définie, - parfois considérée comme une période Proto-Unetice - (Besse et Desideri 2005). L’analyse de la céramique commune a permis de saisir l’importance du substrat régional, en l’occurrence le Cordé, dans son expression. En effet, certains types céramiques du Cordé perdurent au Campaniforme et en représentent la majorité des types céramiques (Besse 2001, p. 69). Le rituel funéraire campaniforme, dans le domaine oriental, s’inscrit dans un complexe évolutif allant du Néolithique final au Bronze ancien. Chaque période possède ses règles d’inhumation, qui sont plus ou moins strictes. Elles sont graduellement moins bien honorées lors des périodes successives (Turek et Peška 2001). Pendant le Campaniforme, les sépultures sont des inhumations individuelles en pleine terre dont la position et l’orientation du corps diffèrent selon le sexe de l’individu. L’homme est orienté nordsud, en décubitus latéral gauche; en revanche la femme est orientée sud-nord, en décubitus latéral droit. Le point commun entre les deux sexes est la position de la tête de l’inhumé, le regard orienté à l’est (Müller A. 1998). On retrouve en Bohême un certain nombre de nécropoles qui sont, en général, constituées d’une vingtaine de tombes seulement. Des éléments cordés et campaniformes se retrouvent parfois dans le même gisement, comme à Čachovice (Mlada Boleslav) et à Brandýsek (Slany), voire dans la même tombe, comme dans le gisement de Sulejovice (Litoměřice) (Moucha 1992). Selon C. Strahm, les rituels 54 Du groupe culturel à l’individu funéraires des cultures Cordé et Campaniforme, qui sont à la fois similaires et divergents, ne peuvent pas s’être développés sans un apport réciproque et doivent avoir coexisté avec une indépendance réciproque (Strahm 1998, p. 36). Par ailleurs, les différences formelles dans la culture matérielle et le rituel funéraire entre le Cordé et le Campaniforme ont lieu, pour certains chercheurs comme J. Turek et J. Peska (2001), à l’intérieur d’un même système symbolique. Selon eux, il est bien probable que l’origine de cette culture soit la conséquence de transformations internes au sein d’une même société. La proportion relative d’habitats par rapport au nombre de sites était de 4 % il y a plus de trente ans, elle est aujourd’hui d’environ 15 % (Turek et Peška 2001). Les structures domestiques vont probablement permettre d’affiner, dans le futur, les connaissances que l’on peut avoir du Campaniforme en Bohême. Les données provenant de l’archéologie ne semblent pas plaider en faveur d’une rupture de peuplement avec l’émergence du Campaniforme. Néanmoins, pour certains chercheurs, le Campaniforme résulterait de migrations depuis le sud-ouest ; pour d’autres, il s’agirait plutôt, d’une transformation soudaine et complexe de la culture Cordé (Zapatocký 1994). 2.2.3. Le Campaniforme en Bohême : les données de l’anthropologie biologique Les données crâniométriques concernant la Bohême s’intègrent parfaitement à l’image que l’on peut avoir des populations de la fin du Néolithique en Europe (Menk 1979, 1981). En effet, les individus cordés présentent une morphologie franchement dolichocéphale ou crâne allongé, contrairement aux individus appartenant à la culture Campaniforme qui possèdent cette forme bien caractéristique arrondie - la brachycéphalie. Au Bronze ancien, cette morphologie arrondie a tendance à s’effacer au profit de formes plus intermédiaires, voire allongées. Une autre étude concernant les variations morphologiques crâniennes de la fin du Néolithique dans cette région a été menée par V. Černý (1996). Il a étudié un échantillon composé de plus de 200 individus provenant des cultures Cordé, Campaniforme, Unetice et Knoviz. Les analyses montrent une proximité au niveau de la morphologie crânienne pour les individus appartenant à la culture Cordé et à celle d’Unetice. Le Campaniforme quant à lui se distingue des deux cultures précédentes et se rapproche de la culture de Knoviz. On retrouve ainsi les mêmes tendances que pour l’analyse de R. Menk (1979, 1981). Malgré cela, V. Černý n’exclut pas complètement une origine autochtone du Campaniforme car les divergences entre groupes culturels sont dues avant tout à des différences du segment neurocranial. Ce dernier, is regarded as having a limited taxonomical value in population genetics studies (Černý 1996, p. 333). Il propose as a working hypothesis for future research, certain autochtonous microevolutionary processes to explain this observed variability (Černý 1996, p. 340). On peut également citer l’étude d’A. Czarnetzki (1984) concernant les caractères discrets crâniens et infracrâniens de ces mêmes populations. Il a calculé des distances entre les hommes et les femmes provenant des cultures Cordé, Campaniforme et Unetice. Selon ses résultats, les caractères discrets prouveraient qu’il y aurait eu des mouvements migratoires des hommes pendant le Campaniforme uniquement. En revanche, les femmes montreraient une morphologie similaire à celles des femmes des deux autres périodes, allant dans le sens d’une stabilité des individus de sexe féminin dans le temps. Une étude basée sur les analyses chimiques des ossements humains ayant comme perspective de déceler d’éventuelles migrations pendant le Campaniforme en Europe centrale a été menée par D. Price et ses collaborateurs (Price et al. 2004)8. Ils ont ainsi mesuré les taux isotopiques du strontium contenus dans les os et l’émail dentaire de six individus provenant de trois nécropoles - deux localisées en Bohême, Kněževes et Velké Přílepy, et une en Moravie, Moravska Nova Ves. Deux individus seulement sur les six analysés semblent d’origine locale. Trois des sujets sont des hommes (dont deux d’origine allochtone) et trois autres indéterminés (dont deux d’origine allochtone). Ces diverses études présentent une même tendance : une mobilité certaine des individus pendant le Campaniforme. Néanmoins, cette mobilité s’interprète différemment selon l’approche utilisée : mobilité de l’homme, d’une partie de la population seulement... 8 Se référer au chapitre 1.5. pour une synthèse des résultats sur l’ensemble de l’aire étudiée (l’Europe centrale). 55 Chapitre 2 2.2.4. Le Campaniforme en Bohême : les échantillons étudiés Pour cette région, les populations analysées appartiennent aux cultures Cordé, Campaniforme et Unetice. Elles se composent de 507 individus répartis sur 64 sites (figure 9). Ceux-ci sont essentiellement localisés dans les parties centrale et septentrionale de la Bohême. Deux sites - Lochenice (n°21) et Plotiště n. Labem (n°39) - se situent dans sa partie orientale. Seul le site de Zelesice (n° 64) comportant 1 individu Campaniforme est situé en Moravie méridionale. Les échantillons ont des effectifs très variables - de 1 à 90 individus. Sur l’ensemble des sites, 24 d’entre eux appartiennent à la culture Cordé, 15 à la culture Campaniforme et 14 au Bronze ancien (culture d’Unetice). Il faut ajouter à ce décompte 11 sites présentant une situation mixte. Seuls deux sites comportent des individus appartenant à l’ensemble des cultures étudiées. Il s’agit de Březno (n°5) et Sulejovice (n°53). Les sites de Čachovice (n°7) et de Radovesice (n°42) sont à la fois cordés et campaniformes. Les sites de Plotiště n. Labem (n°39) et Rvenice I. (n°44) montrent des sépultures datant du Campaniforme et du Bronze ancien. Finalement, les sites de Blšany (n°2), Chotěbudice (n°8), Malé Čičovice (n°23), Pha8-Čimice (n°35) et Vikletice (n°60) présentent des occupations de la culture Cordé et du Bronze ancien. Figure 9 : La Bohême, répartition des sites étudiés. Nous avons attribué à chaque site un numéro reporté sur la carte, puis figurent le nom des sites, leur(s) attribution(s) culturelle(s) et, entre parenthèses, leur effectif. 1. Bílina : Cordé (3) / 2. Blšany : Cordé (1) ; Unetice (11) / 3. Brandýsek : Campaniforme (17) / 4. Břešt’any : Cordé (6) / 5. Březno : Cordé (2) ; Campaniforme (1) ; Unetice (37) / 6. Brodce : Unetice (15) / 7. Čachovice : Cordé (34) ; Campaniforme (5) / 8. Chotěbudice : Cordé (1) ; Unetice (6) / 9. Cítov : Cordé (1) / 10. Dobroměřice : Cordé (1) / 11. Dřevčice : Cordé (1) / 12. Hrdlovka : Cordé (2) / 13. Hrušov : Unetice (5) / 14. Kleneč : Cordé (1) / 15. Klučov : Cordé (4) / 16. Kněževes : Campaniforme, (13) / 17. Konobrže : Cordé (1) / 18. Kozly : Campaniforme (1) / 19. Kučlin : Cordé (1) / 20. Libochovice : Campaniforme (1) / 21. Lochenice : Campaniforme (10) / 22. Malé Březno : Unetice (3) / 23. Malé Čičovice : Cordé (1) ; Unetice (2) / 24. Ml. Bol. Cejeti : Unetice (8) / 25. Mochov : Campaniforme (21) / 26. Most : Cordé (1) / 27. Obrnice : Cordé (4) / 28. Patokryje : Cordé (1) / 29. Pha5-Jinonice : Cordé (6) / 30. Pha5-Malá Ohrada : Unetice (47) / 31. Pha5-Nové Butovice : Unetice (3) / 32. Pha5-Řeporyje : Campaniforme (1) / 33. Pha5-Smíchov : Unetice (6) / 34. Pha8-Bohnice : Unetice (3) / 35. Pha8-Čimice : Cordé (1) ; Unetice (6) / 36. Pha8-Kobylisy : Campaniforme (2) / 37. Pha9-Čakovice : Cordé (2) ; Unetice (10) / 38. Pha9-Kbely : Unetice (3) / 39. Plotiště n. Lab. : Campaniforme (1) ; Unetice (13) / 40. Poláky : Cordé (7) / 41. Prosetice : Cordé (1) / 42. Radovesice : Cordé (3) ; Campaniforme (12) / 43. Rosnice : Campaniforme (1) / 44. Rvenice I. : Campaniforme (1) ; Unetice (5) / 45. Rvenice II : Unetice (1) / 46. Selibice : Campaniforme (1) / 47. Široké Třebčice : Cordé (2) / 48. Skupice-Stracotin : Campaniforme (1) / 49. Skyřice : Cordé (1) / 50. Soběsuky : Cordé (3) / 51. Stadice : Cordé (3) / 52. Stara Kouřim : Cordé (1) / 53. Sulejovice : Cordé (8) ; Campaniforme (1) ; Unetice (4) / 54. Svépravice : Cordé (1) / 55. Trmice : Cordé (11) / 56. Tuchoměřice : Campaniforme (1) / 57. Tursko : Unetice (6) / 58. Tvršice : Unetice (2) / 59. Velké Přílepy : Campaniforme (6) / 60. Vikletice : Cordé (90) ; Unetice (3) / 61. Vraný : Unetice (3) / 62. Žabovřesky : Campaniforme (1) / 63. Zabrušany : Cordé (1) / 64. Zelešice : Campaniforme (1). 56 Du groupe culturel à l’individu 2.3. Le domaine oriental : la Hongrie L’intégration de la Hongrie dans notre étude a un intérêt certain à plus d’un titre. Premièrement, cette région représente l’extension la plus orientale de la manifestation. Ensuite, le Campaniforme hongrois se distingue des autres ensembles d’Europe centrale puisque le substrat n’y est pas le Cordé. Enfin, son occupation se limite à la région entourant Budapest. Ces différents éléments intriguent et suscitent, parmi les différentes aires touchées par le Campaniforme, un intérêt grandissant. 2.3.1. Le Campaniforme en Hongrie : son insertion culturelle, spatiale et temporelle Le Campaniforme hongrois est intégré au groupe de Csepel (Campaniforme-Csepel), attribué à une phase moyenne du Bronze ancien. Les découvertes se concentrent uniquement dans la région de Budapest. Afin de situer le contexte d’apparition de ce phénomène en Hongrie, nous allons survoler les différentes cultures qui constituent le Bronze ancien, ce dernier se subdivisant en trois phases : Le Bronze ancien I est représenté en Hongrie dans ses parties centrale et orientale par la culture de MakóKosihy-Caka - substrat sur lequel le Campaniforme se développe - et par une phase ancienne du groupe Somogyvár-Vinkovci dans sa partie occidentale. Le Bronze ancien II voit non seulement la perduration des cultures de Makó-Kosihy-Caka, dont l’aire se restreint à la partie la plus septentrionale de la Hongrie, et de Somogyvár-Vinkovci dans sa partie occidentale, mais également l’émergence d’une série de groupes culturels. Ainsi, le groupe du Campaniforme-Csepel se développe dans la région de Budapest, le Nagyrev ancien se développe dans la partie centrale de la Hongrie, le groupe d’Obedà-Pitvaros dans sa partie méridionale et celui de Nyirseg dans sa partie orientale. Pendant le Bronze ancien III, des différents groupes culturels présents lors de la phase précédente, seul celui de Nagyrev persiste. A côté de cela, de nouveaux groupes se développent comme la culture de Perjamos dans l’aire méridionale, celles de Szaniszlo et d’Ottomany dans l’aire orientale, de Kisapostag dans la partie occidentale et d’Havtan dans le nord de la Hongrie. Le Bronze ancien s’étend de 2600-2500 à 2000-1900 av. J.-C. (Kalicz-Schreiber et Kalicz 2001). Nous avons vu lors du chapitre précédent que les datations du Campaniforme-Csepel surprennent puisqu’une série de dates anciennes place cette manifestation dans la première moitié du 3e millénaire et que de nouvelles datations la situent dans la seconde moitié, ce qui est un meilleur reflet de l’occupation temporelle du phénomène dans sa partie orientale (Vander Linden 2006). 2.3.2. Le Campaniforme en Hongrie : les données de l’archéologie Les gisements se situent tout le long du Danube dans la région de Budapest avec une concentration de sites très élevée dans l’île de Csepel, qui se monte à plus de 60 sites (Endrödi 2003). Le rituel funéraire du groupe de Makó est l’incinération. L’inhumation semble advenir avec le Campaniforme. En effet, les sépultures campaniformes sont représentées tant par des incinérations que par des inhumations. Les nécropoles peuvent contenir un nombre élevé d’individus. La nécropole de Bekasmegyer (Budapest) présente notamment plus d’une centaine de sépultures dont la majorité sont des incinérations en urne et en fosse, mais on trouve aussi des inhumations (Kalicz-Schreiber et Kalicz 2001). Les structures funéraires se situent principalement à proximité des habitats comme à Szigetszentmiklòs (Endrödi 1998) ou à Albertsfalva (Endrödi 2003). L’arrangement des sépultures est parfois peu ordinaire, comme sur le gisement de Tököl où les inhumations sont alignées sur près d’un kilomètre (Vander Linden 2006). Le matériel présente des grandes affinités, non seulement avec la culture de Nagyrev dans sa phase ancienne qui est contemporaine au Campaniforme-Csepel, mais également avec les cultures qui la 57 Chapitre 2 précèdent, soit les groupes de Makó-Kosihy-Caka et Somogyvár-Vinkovci (Endrödi 1998). Les vestiges caractérisant le Campaniforme, comme la céramique décorée, pointes de flèche ou encore boutons perforés en V... se retrouvent en contexte funéraire uniquement dans les inhumations ou dans les sépultures à cendres dispersées (Endrödi 2003). Les structures domestiques sont abondantes pour cette région. Leur architecture est parfois comparable à celle du groupe du Nagyrev ancien (Vander Linden 2006). A la fin de la période de Makó-Kosihy-Caka, il est possible de mettre en évidence une infiltration campaniforme depuis l’ouest (Endrödi 1998). Pour A. Endrödi (2003), le Campaniforme-Csepel serait issu d’influences, voire d’arrivées de petits groupes, provenant de la région de la République Tchèque. Selon certains chercheurs, les données archéologiques prouveraient que les campaniformes hongrois ont vécu dans un territoire limité dans l’espace sur une courte période et qu’ils ne se seraient pas mélangés aux populations locales (Zoffmann 2000). Pour d’autres, puisqu’une partie des vestiges campaniformes montre des associations avec la culture de Nagyrev, on ne peut nier l’hypothèse de contacts entre les différentes cultures (Endrödi 1998). Ainsi, en considérant ses caractéristiques culturelles, le Campaniforme hongrois est un groupe culturel autonome ayant eu des contacts plus ou moins intenses avec ses groupes voisins (Endrödi 1998). R. Kalicz-Schreiber et N. Kalicz (2001), quant à eux, estiment que l’on ne doit pas considérer le Campaniforme-Csepel comme une culture indépendante. Cette manifestation doit être interprétée comme un reflet de contacts au sein de la sphère sociale constituée de plusieurs cultures et territoires distincts. De plus, elle devrait être intégrée dans une phase ancienne de la culture de Nagyrev et son apparition serait liée à des influences méridionales (Kalicz-Schreiber et Kalicz 2001). Le Campaniforme hongrois intrigue et les interprétations issues des données provenant de l’archéologie ne s’accordent pas réellement. De l’apport de sang nouveau au développement interne de populations locales, il semble difficile de trancher. 2.3.3. Le Campaniforme en Hongrie : les données de l’anthropologie biologique Si le Campaniforme hongrois est abondamment représenté par des structures domestiques, celles funéraires sont moindres. Il s’agit principalement d’incinérations et les quelques inhumations fournissent des informations très lacunaires. Une étude menée par Z.K. Zoffmann (2000) a concerné un bilan des recherches en paléoanthropologie des populations préhistoriques dans le Bassin des Carpates. Nous n’avons pas d’informations sur les études qui ont été réalisées sur les individus campaniformes, mais selon l’auteure, although there are very few evaluable anthropological finds from the biritual cemeteries of the Bell-beakers in the Carpathian Basin, the appearance of the characteristic planoccipital Taurid type, unknown until then from the Carpathian Basin, in the populations of some later cultures [...] suggests a mixture with the local population against the archaeological theories (Zoffmann 2000, p. 76-77). D. Price et ses collaborateurs (2004)9, dans leur étude concernant l’analyse des taux isotopiques du strontium des individus campaniformes en Europe centrale, ont intégré six sujets hongrois, sans attribution sexuelle, provenant de trois sites localisés près de Budapest (Bekasmegyer, Csepel II et Szigetszentmiklòs). Selon leurs résultats, cinq des six sujets analysés semblent d’origine allochtone. Les données provenant de l’étude des individus rattachés au Campaniforme hongrois ne sont pas nombreuses, elles fournissent néanmoins des éléments intéressants. Il semblerait qu’il y ait un apport de sang nouveau lors du Campaniforme, lequel n’exclut pas la présence de contacts avec les populations voisines, ce qui a été suggéré par la morphologie crânienne. Cependant, les effectifs sont petits et les informations concernant certaines études sont éparses, ainsi une prudence particulière quant aux interprétations est nécessaire. 9 58 Se référer au chapitre 1.5. pour une synthèse des résultats sur l’ensemble de l’aire étudiée (l’Europe centrale). Du groupe culturel à l’individu 2.3.4. Le Campaniforme en Hongrie : les échantillons étudiés Pour la Hongrie, les populations analysées appartiennent aux deuxième (Bronze ancien II) et troisième (Bronze ancien III) phases du Bronze ancien. Elles se composent de 92 individus répartis sur 15 sites (figure 10). Le substrat du Campaniforme (Makó-Kosihy-Caka) n’est malheureusement pas représenté ici. Les sites intégrés dans ce travail sont essentiellement localisés dans les parties centro-septentrionale (région autour de Budapest) et méridionale (région autour de Szeged). Trois ensembles se situent en dehors de ces zones. Le site de Kaposvàr elkenülo (n°7) appartenant à la culture Somogyvar Vinkovci se trouve dans la partie occidentale du pays. Les échantillons de Rákóczifalva (n°11) - culture de Nagyrev et de Tiszakeszi (n°15) - culture de Nyirseg -, se situent dans la partie orientale de la Hongrie. Les effectifs sont très variables - de 1 à 24 individus. Sur l’ensemble des sites, les échantillons se répartissent de la manière suivante : - Le Bronze ancien II est représenté par les ensembles campaniformes, soit les sites d’Albertfalva (n°1), de Bekasmegyer (n°2), de Csepel-Vizmä (n°3) et de Szigetszentmiklòs (n°13). Mais aussi des sites des cultures d’Obedà-Pitvaros comme Homokbànya (n°6), Kiskundorozsma (n°8) et Sándorfalva-Eperjes (n°12), de Nyirseg avec le site de Tiszakeszi (n°15) et de Somogyvar Vinkovci avec le gisement de Kaposvàr elkenülo (n°7). - Le Bronze ancien III est composé par les sites de Deszk A (n°4), de Dunakeszi Szedesdoto (n°5), de Pitvaros (n°10) et de Rákóczifalva (n°11) appartenant à la culture de Nagyrev et par les sites de Hòdmezövàsàrhely-Kopànos (n°9) et de Szöreg (n°14) appartenant à la culture de Perjamos. Figure 10 : La Hongrie, répartition des sites étudiés. Nous avons attribué à chaque site un numéro reporté sur la carte, puis figurent le nom des sites, leur attribution culturelle et, entre parenthèses, leur effectif. 1. Albertfalva : Campaniforme-Csepel (1) / 2. Bekasmegyer : Campaniforme-Csepel (14) / 3. Csepel-Vizmä : Campaniforme-Csepel (3) / 4. Deszk A : Nagyrev (14) / 5. Dunakeszi Szedesdoto : Nagyrev (1) / 6. Homokbànya : Obedà-Pitvaros (1) / 7. Kaposvàr elkenülo : Somogyvar Vinkovci (2) / 8. Kiskundorozsma : Obedà-Pitvaros (4) / 9. Hòdmezövàsàrhely-Kopànos : Perjamos (2) / 10. Pitvaros : Nagyrev (19) / 11. Rákóczifalva : Nagyrev (1) / 12. Sándorfalva-Eperjes: Obedà-Pitvaros (5) / 13. Szigetszentmiklòs : Campaniforme-Csepel (1) / 14. Szöreg : Perjamos (24) / 15. Tiszakeszi : Nyirseg (2). 59 Chapitre 2 2.4. Le domaine méridional : le sud de la France Les connaissances que l’on a du Campaniforme en France ne sont pas comparables sur l’ensemble du territoire. La densité et le type de sites montrent de grandes inégalités. Ainsi, certaines régions sont abondamment documentées, comme le sud de la France ou encore la Bretagne. C’est pourquoi, nous avons choisi la première de ces deux régions pour tenter d’y appréhender les modalités d’apparition du Campaniforme. Malgré l’abondance des données, la fin du Néolithique ici se révèle difficile à aborder. 2.4.1. Le Campaniforme dans le sud de la France : son insertion culturelle, spatiale et temporelle La fin du Néolithique coïncide avec l’émergence d’une série de groupes culturels délimités dans l’espace lors de la désarticulation de l’unité chasséenne qui prévalait jusqu’alors sur une grande partie du territoire. Ainsi, le sud de la France voit l’apparition de groupes culturels tels le Saint-Ponien (3500-3000 av. J.-C.) et le Vérazien (3000-2500 av. J.-C.) dans le Languedoc occidental, celui de Ferrières (32002800 av. J.-C.) et le Fontbouisse (2800-2200 av. J.-C.) dans le Languedoc oriental, de Treilles (3500-2200 av. J.-C.) dans la région des Grands Causses de l’Aveyron et ses marges et un regroupement de différentes unités culturelles, le groupe de Fraischamps (3300-2900 av. J.-C.), le Couronnien (3100-2400 av. J.-C.) et le Rhône-Ouvèze (2800-2200 av. J.-C.), réunies sous le terme de Chalcolithique provençal, bien entendu localisé en Provence. Nous ne nous attarderons pas sur le débat concernant la dénomination de ces différents groupes culturels, néanmoins des problèmes de terminologie se posent puisque certaines des cultures entrant en jeu sont attribuées au Néolithique et d’autres au Chalcolithique (D’Anna 1995, p. 300). Pour J. Guilaine et al. (2001), le terme de Néolithique final devrait être réservé aux cultures de transition entre le 4e et le 3e millénaires, comme le Saint-Ponien, Treilles ou Fraischamps, possédant une métallurgie débutante, et celui de Chalcolithique pour la phase suivante composée des groupes culturels connaissant une métallurgie du cuivre effective, comme le Fontbouisse, le Vérazien, le Couronnien et le Rhône-Ouvèze, selon une terminologie définie lors du colloque de Narbonne en 1970 (Guilaine et al. 2001). Dans ce contexte, le Campaniforme, précédant l’âge du Bronze ancien, se développe lors de la seconde phase. Son insertion au sein du contexte local et les relations chronologiques entre Campaniforme et cultures locales du Néolithique final ont fait l’objet de nombreuses interprétations contradictoires (Lemercier 2002, p. 26), tant la situation est complexe. La répartition du Campaniforme n’est pas uniforme sur l’ensemble de ce territoire. En effet, le sud-est avec plus de 320 sites est de loin la région la plus riche en gisements (Lemercier 2002). Le Campaniforme du sud de la France possède une quarantaine de datations radiocarbone qui le placent dans la première moitié du 3e millénaire (Bailly et Salanova 1999). 2.4.2. Le Campaniforme dans le sud de la France : les données de l’archéologie Les céramiques décorées sont composées à la fois par le standard, défini par L. Salanova (2000, 2002), et par des productions dites locales comme le Rhodano-Provençal et le Pyrénéen. Une périodisation du Campaniforme en quatre phases a été établie sur la base d’une interprétation chronologique des différents horizons de cette céramique (Guilaine et al. 2001). Celle-ci est composée par une phase ancienne constituée presque exclusivement de céramique décorée - le standard - et de structures funéraires. La deuxième, considérée comme une phase de transition ou d’intégration, voit le développement de structures domestiques où la céramique décorée cohabite avec la céramique domestique des diverses traditions locales. Lors de la troisième, on assiste à une diversification des formes céramiques et à l’apparition des traditions céramiques campaniformes locales citées plus haut. Et la dernière phase correspond au Campaniforme barbelé (Lemercier 2003) constitué de nouvelles formes céramiques du même nom qui marque la fin du phénomène dans cette région ou le début du Bronze ancien pour certains chercheurs. 60 Du groupe culturel à l’individu En ce qui concerne la céramique domestique, les types [...] sont nouveaux, peu d’entre eux trouvent une origine dans les cultures du substrat régional de la fin du Néolithique. De plus, les sites avec céramiques décorées sont bien plus nombreux que les sites avec céramique commune seule, renforçant le renouvellement des composantes culturelles au Campaniforme (Besse 2001, p. 69). Nous avons vu qu’il est possible aujourd’hui de mettre en évidence, par l’étude de la céramique décorée (Lemercier 2002) et par celle de l’industrie lithique (Furestier 2005), deux phases distinctes dans l’évolution du Campaniforme, du moins, dans le sud-est de la France. Une phase ancienne et hermétique d’apparition du phénomène et une seconde d’intégration et diffusion au sein des traditions locales. Alors que le midi de la France et, dans une moindre mesure, le centre-ouest connaissent une métallurgie bien développée durant la première moitié du 3e millénaire, les cultures les plus septentrionales [...] sont elles restées, volontairement ou non, en marge des innovations technologiques (Vander Linden 2006, p. 83). Toutefois, pendant le Campaniforme, les marques de métallurgie ne sont pas des plus abondantes (Vander Linden 2006). Les divers types de sépultures campaniformes rencontrées dans le sud de la France ont été présentés lors du chapitre précédent. Le rituel le plus fréquent est ainsi la réutilisation d’inhumations collectives qui peuvent être des dolmens, des hypogées ou encore des grottes sépulcrales datant du Néolithique finalChalcolithique. Quelques sépultures individuelles strictement campaniformes peuvent tout de même être mises en évidence. Nous avons également vu que le Campaniforme est documenté dans cette région par un nombre important de structures domestiques. Les sites sont soit « purs », présentant uniquement du matériel attribué au Campaniforme, soit « mixtes » regroupant des éléments appartenant aussi bien au Campaniforme qu’aux diverses cultures locales de la fin du Néolithique. Les données archéologiques marquent, dans une phase initiale de développement du Campaniforme, une rupture certaine de ce dernier avec les cultures du Néolithique final local. La rupture est suivie par une cohabitation ou intégration plus ou moins marquée selon les vestiges étudiés. Pour O. Lemercier (2004, p. 235), la diffusion du Campaniforme pourrait avoir eu lieu par voie maritime depuis le Portugal concernant des petits groupes porteurs à la fois d’objets mais surtout de savoir-faire. 2.4.3. Le Campaniforme dans le sud de la France : les données de l’anthropologie biologique Les analyses anthropologiques dans une perspective du peuplement campaniforme reposent ici encore et seulement sur les données métriques crâniennes. Les premières études ne constatent pas la fameuse brachycéphalie planoccipitale caractérisant les populations campaniformes de l’est de l’Europe. Les individus sont franchement dolichocéphales poursuivant la morphologie locale méditerranéenne. Quelques exceptions peuvent être mises en évidence au sein d’ossuaires campaniformes réutilisés pendant le Bronze ancien (Riquet et al. 1963). R. Menk (1979, 1981) a également inclus le sud de la France dans son étude. Il a analysé 78 individus de la fin du Néolithique au Bronze ancien, dont vingt-trois campaniformes. Pour cet auteur, la grande majorité des individus campaniformes se regroupe parmi les brachymorphes ou les méso-dolichomorphes à grandes dimensions se distinguant ainsi du fonds local (dolichomorphes). Les individus du Bronze ancien, quant à eux, se partagent entre ces différentes morphologies. Nous avons tout de même vu lors du chapitre précédent que des réévaluations de corpus ancien placent aujourd’hui certains individus brachycéphales dans le Bronze ancien plutôt qu’au Campaniforme (Tchérémissinoff 2006). La question reste donc ouverte. Au-delà de cela, très peu d’études ont concerné les individus campaniformes. Le contexte confus des structures funéraires présentant plusieurs occupations dans lesquelles il n’est pas toujours aisé de distinguer les Campaniformes avec certitude en est vraisemblablement la cause. 61 Chapitre 2 2.4.4. Le Campaniforme dans le sud de la France : les échantillons étudiés La situation pour le sud de la France n’est pas des plus aisées. La pratique funéraire usuelle au Campaniforme est la réutilisation des sépultures collectives de la fin du Néolithique. Celles-ci contiennent ainsi plusieurs occupations qui sont parfois difficiles à distinguer. Les populations étudiées ici, constituées de près de 700 individus et de 5 sites, se répartissent entre le Néolithique final/Chalcolithique et le Bronze ancien (figure 11). Celles-ci sont principalement localisées dans le sud-est de la France. Seul le dolmen des Peirières (n°1) se situe dans la partie occidentale de l’aire considérée. Les effectifs varient entre 18 et 172 individus. Les ensembles sont complexes d’un point de vue de leur attribution culturelle, ainsi nous les présentons ici par monument : - Le dolmen des Peirières (n°1) possède deux phases culturelles distinctes, la première datant de la fin du Néolithique et la seconde du Campaniforme. - Le dolmen des Peyraoutes (n°2) réunit également deux occupations, dont l’une (couche 2- C2) pourrait être campaniforme et l’autre serait chalcolithique (couche 3 - C3). Malheureusement le remaniement des couches (Bouville 1971, Lemercier 2002), nous amène à travailler avec un unique groupe Campaniforme/Chalcolithique sans les distinguer. - L’hypogée des Crottes (n°3) présente deux occupations chalcolithiques : couche 2 (C 2) et la couche 5 (C5). La première (C2) est-elle campaniforme ? Nous testerons cette hypothèse lors des analyses. - L’hypogée des Boileau (n°4), quant à lui, est à rattacher au Néolithique final. - Le dolmen des Cudières (n°5) présenterait deux phases d’occupation - Chalcolithique et Bronze ancien qu’on ne saurait différencier et qui forment ici un seul ensemble. Figure 11 : Le sud de la France, répartition des sites étudiés. Nous avons attribué à chaque site un numéro reporté sur la carte, puis figurent le nom des sites, leur(s) attribution(s) culturelle(s) et, entre parenthèses, leur effectif. 1. Dolmen des Peirières : Néolithique final (18) ; Campaniforme (70) ; indéterminé (19) / 2. Dolmen des Peyraoutes : Campaniforme/ Chalcolithique (172) / 3. Hypogée des Crottes : Chalcolithique couche 2 (49) ; Chalcolithique couche 5 (92) / 4. Hypogée des Boileau : Néolithique final (130) / 5. Dolmen I des Cudières-Saint Bacchi : Chalcolithique/Bronze ancien (98). 62 Du groupe culturel à l’individu 2.5. Le domaine méridional : le nord de l’Espagne La péninsule Ibérique, au même titre que la Bohême, est une région incontournable dans la compréhension du phénomène Campaniforme. Elle a été retenue à maintes reprises comme un foyer d’apparition possible de ce complexe culturel. Nous avons choisi d’orienter notre recherche sur le nord de l’Espagne et plus précisément la Meseta septentrionale et centrale puisqu’il s’agit d’une région très bien documentée. 2.5.1. Le Campaniforme dans le nord de l’Espagne : son insertion culturelle, spatiale et temporelle En Meseta, les premières données néolithiques se placent lors de la moitié du 4e millénaire avant notre ère. Le Chalcolithique prend place au début du 3e millénaire. Le Campaniforme, quant à lui, apparaît dans la phase récente de l’âge du Cuivre et coexiste avec lui jusqu’à l’émergence du Bronze ancien. On sait aujourd’hui, grâce à ces vingt dernières années de recherche, que le phénomène campaniforme se développe sur l’ensemble de la région. Aujourd’hui, les découvertes sont importantes et les vestiges sont largement représentés par les structures domestiques. Dans la Meseta, le Chalcolithique précampaniforme se développe dans la première moitié du 3e millénaire et se termine vers 2200 av. J.-C. Les datations radiocarbone placent le début du Campaniforme dans la première moitié du 3e millénaire avant notre ère en péninsule Ibérique, mais elles se situent entre 2500 et 2200 av. J.-C. dans la Meseta. Il voit le développement d’un horizon local, le Ciempozuelos, occupant sa phase finale et perdurant jusqu’au Bronze ancien (Garrido-Pena 2000). 2.5.2. Le Campaniforme dans le nord de l’Espagne : les données de l’archéologie La céramique décorée est constituée à la fois des gobelets maritimes, que l’on retrouve principalement dans le sud de la Meseta et davantage en contexte funéraire, et de la céramique de type Ciempozuelos, très stylisée de couleur noire ou grise avec décoration incisée et constituée avant tout de petits bols (Vazquez Cuesta 2006). Celle-ci est de loin la céramique la plus abondante dans les gisements. Elle concerne, en effet, plus de 87% de la céramique retrouvée (Garrido-Pena et al. 2005). Pour R. Harrison (1986), une phase ancienne du Campaniforme peut être mise en évidence par le premier type de céramique et ses dérivés. Celle-ci est suivie par une phase récente composée par la céramique de l’horizon local - le Ciempozuelos. Cette seconde phase a une durée plus longue (Harrison 1986). Une étude de plus de 200 céramiques décorées de la Meseta centrale a été réalisée par R. Garrido-Pena (2000). Il a relevé, d’une part, une standardisation tant au niveau des dimensions que de la capacité volumétrique de certaines céramiques, et de l’autre, d’après l’étude des décors, des réseaux d’échanges à plusieurs niveaux, - local, régional et interrégional (Garrido-Pena et al. 2005). Ces résultats, selon l’auteur, confirment le statut de ces céramiques comme des objets à haute valeur sociale et permet de rejeter celui d’objets ordinaires ou domestiques (Garrido-Pena 2000). Les études pétrographiques montrent que les céramiques sont produites localement et que la proportion des dégraissants varie selon les gisements (Millàn et Arribas 1994). Des études chimiques de céramiques des sites de la vallée de Ambrona, tels le tumulus de la Sima ou encore de la Pena de Abuela, ont montré qu’une partie d’entre elles ont contenu de la bière (Garrido-Pena et al. 2005). Cet élément permet de mettre à nouveau sur les rangs la relation entre céramique campaniforme et contenant rituel (Garrido-Peno 2000). Pour d’autres, the bell beaker beer is not a novelty [...] we can not assume without criticism that beer is a high value product in bell beaker time (Vazquez Cuesta 2005, p. 87). 63 Chapitre 2 La métallurgie campaniforme suit les traditions technologiques de ses prédécesseurs (Vander Linden 2006) et les aires d’approvisionnement du minerai pourraient se localiser dans les régions de Madrid et de Toledo (Garrido-Pena et al. 2005). Les structures domestiques se situent en grande partie près de points d’eau et en hauteur with an ample environment control (Garrido-Pena et al. 2005). Ces sites présentent fréquemment une durée d’occupation longue à travers le temps et les différentes superpositions rendent les analyses de ces structures difficiles (Garrido-Pena 2000). La Meseta s’intègre parfaitement à la diversité occidentale offerte par le Campaniforme à travers son rituel funéraire. Ainsi, de la réutilisation de monuments mégalithiques néolithiques ou chalcolithiques précampaniformes, aux inhumations sous tumulus ou en grotte, à la tombe plate en pleine terre, pour ne citer que quelques exemples, la variabilité est grande contrairement à l’unité de la sépulture collective durant le Néolithique (Garrido-Pena 2000). La disposition de l’inhumé campaniforme, ainsi que le matériel qui l’accompagne, au sein des sépultures en réemploi, est souvent discutée et considérée comme un traitement particulier du défunt ayant une signification sociale (Garrido-Pena et al. 2005). Ces données archéologiques conduisent R. Garrido-Pena, M. Rojo Guerra et I. Garcia Martinez De Lagran (2005) à considérer les interprétations fonctionnelles par l’échange de biens socialement valorisés - et particulièrement les postulats de D.L. Clarke (1976) - comme une explication possible - voire probable - du développement du Campaniforme dans la Meseta. Ainsi, les faits provenant de l’archéologie dans cette région vont dans le sens de l’hypothèse selon laquelle Bell Beakers and accompanying objects are not vulgar domestic elements, but high value objects used in the legitimation strategies of the leaders emerged in the Meseta Copper Age societies, in the context of a social and economic transformation process, which would have extended through the dense exchange networks, in diverse social mechanisms such as the gift exchanges, politics pacts, marriage exchanges, and so on (Garrido-Pena et al. 2005, p. 450). 2.5.3. Le Campaniforme dans le nord de l’Espagne : les données de l’anthropologie biologique Dans son analyse, R. Menk (1979, 1981) dispose de douze individus provenant essentiellement de Catalogne. Seuls quatre d’entre eux sont franchement brachycéphales et se détachent du fonds néolithique local de manière évidente. Ceux-ci sont considérés par l’auteur comme des individus clairement intrusifs. En revanche, M.D. Garralda (1979), qui a effectué une synthèse concernant la morphologie crânienne des populations du Néolithique et du Bronze ancien en péninsule Ibérique, montre que la composition morphologique varie peu d’une région à l’autre avec une prédominance nette du type méditerranéen gracile (dolichocéphale). Les rares cas de brachycéphalie sont souvent considérés comme à la limite de la variabilité locale. La morphologie arrondie de quelques individus, découverts dans le littoral atlantique, la Catalogne et à Majorque, est mise en parallèle avec celle alpine ou dinarique mais sans aucune certitude (Garralda 1979). Au-delà des études anthropologiques élémentaires concernant l’identification biologique et le recrutement funéraire des individus inhumés (Gonzalez Martin et Campo Martin 2002, Alt 2005, Nicklisch et al. 2005), quelques études ont touché d’autres aspects de la caractérisation de ces populations. Des analyses des isotopes (Trancho et al. 1996, Trancho et Robledo 1999, Dürrwächter et al. 2005) ont été réalisées dans une perspective de détermination de la paléonutrition de quelques ensembles chalcolithiques avec ou sans Campaniforme et du Bronze ancien. Ces études ont montré un type d’économie basée sur une alimentation plutôt végétale avec un léger apport en protéines animales et ce, malgré une faiblesse d’échantillons et de grands problèmes de diagenèse. Une tentative d’analyse de l’ADN des individus de deux gisements situés dans la Valle de Ambrona (le Tumulo de la Sima et la Peña de Abuela) a été réalisée, sans succès, puisque aucune extraction n’a pu être effectuée (Dürrwächter et al. 2005). D’un point de vue purement anthropologique, aucun élément ne semble défendre un apport de sang nouveau lors de l’émergence du Campaniforme dans la Meseta. Quoi qu’il en soit, les études en péninsule Ibérique ne les recherchent pas non plus... 64 Du groupe culturel à l’individu 2.5.4. Le Campaniforme dans le nord de l’Espagne : les échantillons étudiés Pour cette région, les populations analysées se rapportent au Néolithique final, au Chalcolithique, au Campaniforme et à l’âge du Bronze. Elles s’élèvent à près de 180 individus répartis sur 14 sites (figure 12). Ceux-ci sont essentiellement localisés dans la partie septentrionale de la Meseta. Seul le site de la Valle de las Higueras (n°13) se situe dans l’aire méridionale de la région. Les effectifs sont variables - 1 à 50 individus. Sur l’ensemble des sites, les échantillons se distribuent de la manière suivante : - Le Néolithique final est constitué d’individus provenant de sépultures collectives, telles la Sima (n°8), La Velilla (n°9) et Los Zumacales (n°12). - Le Chalcolithique est représenté par les sépultures des sites de Cerro de la Cabeza (n°2), El Tomillar (n°6), Los Cercados (n°11), Valle de las Higueras (n°13), Villimar (n°14). - Le Campaniforme, quant à lui, est composé des inhumations de la Cueva del Destete (n°3), de Fuente Olmedo (n°7), de la Sima (n°8), et de la Valle de las Higueras (n°13). - Quelques individus appartiennent à l’âge du Bronze. Deux sites - Cueva Rubia (n°4) et Cueva Tino (n°5) - sont à rattacher soit au Bronze ancien, soit au Bronze moyen. Le site de Carrelasvegas (n°1) est clairement attribué à la phase moyenne de l’âge du Bronze. Le faible effectif pour cette période ne permettra probablement pas d’intégrer ces sites dans les analyses. Las Arnillas (n°10) présente deux occupations funéraires - Chalcolithique et Campaniforme. Les vestiges humains des deux occupations ne se discernent pas, c’est pourquoi nous considérerons ce site comme un tout. Figure 12 : Le nord de l’Espagne, répartition des sites étudiés. Nous avons attribué à chaque site un numéro reporté sur la carte, puis figurent le nom des sites, leur(s) attribution(s) culturelle(s) et, entre parenthèses, leur effectif. 1. Carrelasvegas : Bronze moyen (1) / 2. Cerro de la Cabeza : Chalcolithique (9) / 3. Cueva del Destete : Campaniforme (5) / 4. Cueva Rubia : Bronze ancien ou moyen (1) / 5. Cueva Tino : Bronze ancien ou moyen (3) / 6. El Tomillar : Chalcolithique (3) / 7. Fuente Olmedo : Campaniforme (1) / 8. La Sima : Néolithique final (22) ; Campaniforme (3) / 9. La Velilla : Néolithique final (50) / 10. Las Arnillas : Chalcolithique/Campaniforme (24) / 11. Los Cercados : Chalcolithique (1) / 12. Los Zumacales : Néolithique final (18) / 13. Valle de las Higueras : Chalcolithique (tumulus 8, cuevas 30) ; Campaniforme (9) / 14. Villimar : Chalcolithique (1). 65 Chapitre 2 2.6. A l’articulation entre les deux domaines : la Suisse La Suisse se situe géographiquement entre deux grands domaines, - méridional et oriental -, et oppose ainsi ces deux grands courants, non seulement pour le Campaniforme, mais également pour l’ensemble des périodes que nous intégrons dans notre travail. De plus, les découvertes, et, par conséquent, les connaissances que l’on peut avoir du Néolithique, sont inégalement réparties sur l’ensemble du territoire helvétique. 2.6.1. Le Campaniforme en Suisse : son insertion culturelle, spatiale et temporelle L’aire chronologique considérée ici s’étend du 5e au 3e millénaire avant notre ère, soit du Néolithique moyen au Bronze ancien. Une tripartition de la Suisse (occidentale, septentrionale et orientale) doit être réalisée pour présenter rapidement son évolution culturelle à travers le temps. En Suisse occidentale, la succession culturelle précampaniforme à travers le Néolithique se fait par les cultures de Cortaillod, Horgen et Lüscherz/Auvernier Cordé (Néolithique final valaisan pour le Valais). En Suisse septentrionale, elle se compose successivement des cultures Egolzwill, Cortaillod, Horgen et Cordé. En Suisse orientale, celle-ci est représentée par les cultures de Rössen, Pfyn, Horgen et Cordé. Le Campaniforme helvétique se place donc à l’articulation entre la fin du Néolithique et le début du Bronze ancien. Il succède aux cultures d’Auvernier Cordé-Lüscherz et le Néolithique final valaisan - à l’ouest -, et le Cordé - au centre et à l’est. Les découvertes campaniformes dépendent de l’état de la recherche et, à quelques exceptions près, les gisements se situent, pour la plupart, dans les parties occidentale et septentrionale du territoire helvétique. Elles se situent dans la 2e moitié du 3e millénaire avant notre ère entre 2500 et 2200 av. J.-C. 2.6.2. Le Campaniforme en Suisse : les données de l’archéologie Le rituel funéraire est bien connu en Suisse occidentale. Ainsi, le Néolithique moyen est caractérisé par les inhumations de type Chamblandes. Une première occupation (4800 et 4300 av. J.-C.) est composée de nécropoles contenant essentiellement des sépultures individuelles. Il est possible de mettre en évidence des changements du rituel funéraire qui ont lieu vers la seconde moitié du 5e millénaire. Cette seconde phase (4300 et 3300 av. J.-C.) est marquée par l’apparition de sépultures collectives dans les cistes Chamblandes. Les sépultures peuvent être doubles ou multiples, simultanées ou successives. Ces sépultures constituent les éléments annonciateurs de la collectivisation des tombes qui se développe à la fin du Néolithique avec l’apparition des tombes mégalithiques (Moinat et Gallay 1998). Ainsi, la pratique la plus répandue pendant le Néolithique récent et le Campaniforme est la sépulture collective. Malgré des lacunes, il est possible de suivre en Suisse septentrionale cette même évolution. La partie orientale de la Suisse reste à l’écart des sépultures mégalithiques. Quelques inhumations individuelles en pleine terre campaniformes suivant la tradition rencontrée dans le domaine oriental ont été mises au jour (Moinat et Stöckli 1995). Dès le Bronze ancien, on assiste à la fois à la reprise de l’inhumation individuelle, de sépultures au sein de ciste et l’apparition de tumuli. Les structures d’habitats précampaniformes sont les très célèbres stations lacustres suisses. Aucun habitat n’est clairement attesté en zone terrestre ou en hauteur (Hasenfratz et Gross-Klee 1995). En revanche, les structures domestiques campaniformes sont connues dans la portion helvétique occidentale seulement. Il s’agit principalement de sites terrestres, néanmoins de rares découvertes en contexte lacustre peuvent être mises en évidence (Besse 2006). Le Campaniforme helvétique se constituerait, d’après les données archéologiques, à la suite d’influences plus ou moins importantes provenant à la fois de la Méditerranée et d’Europe centrale (Gallay 2006). Pour A. Gallay (2006), les influences méridionales résulteraient d’un phénomène idéologique, tandis que celles orientales dériveraient de mouvements de populations. 66 Du groupe culturel à l’individu 2.6.3. Le Campaniforme en Suisse : les données de l’anthropologie biologique Les résultats des analyses concernant les données métriques pour la Suisse semblent s’intégrer à l’image des populations du Néolithique européen. En effet, les individus du Néolithique moyen possèdent une morphologie franchement dolichocéphale (Menk 1979, 1981). Les individus appartenant à la culture Campaniforme, quant à eux, possèdent une morphologie spécifique ; ils sont ainsi extrêmement brachycrânes, à aplatissement occipital et totalement différents du substrat local (Menk 1979, p. 272). En revanche, les ensembles du Néolithique récent, la période de transition, ne sont pas intégrés dans les analyses puisque les données pour cette période sont insuffisantes. Pour remédier à ce hiatus chronologique dans ses interprétations, R. Menk pose l’hypothèse que le type physique observé pendant le Néolithique moyen perdure jusqu’au Campaniforme, moment où l’on observe une rupture du peuplement. Au Bronze ancien la morphologie arrondie tend à s’effacer au profit de formes plus intermédiaires (mésocrânes) (Simon et Kaufmann 1998). En 1996, une nouvelle étude incorporant un ensemble découvert à la fin des années 1980, - le dolmen MXII du site du Petit-Chasseur à Sion - permet d’infirmer, les déductions de R. Menk (Eades 1996). La rupture nette de la morphologie crânienne avec l’apparition du Campaniforme n’est plus envisageable selon l’auteure, puisque la morphologie des individus du Néolithique récent est de forme plutôt intermédiaire (mésocrânes). Une réévaluation des données métriques crâniennes a été effectuée récemment (Desideri à paraître). Elle permet de nuancer les résultats obtenus par ces différentes études. On passe effectivement de formes très allongées au Néolithique moyen à des formes très arrondies au Campaniforme. On doit cependant modérer les résultats puisqu’on trouve déjà en abondance la forme intermédiaire, la mésocrânie au Néolithique moyen et la majorité des individus sont brachycrânes pendant le Néolithique récent. En dépit des effectifs relativement réduits, il est tout de même possible de considérer la présence d’une évolution graduelle de la morphologie crânienne pendant le Néolithique, mais la brachycéphalisation ne semble pas étroitement liée à l’émergence de la culture Campaniforme (Desideri à paraître). Deux études ont été réalisées sur les variations non métriques du crâne et de la dentition (Eades 1996, Desideri 2001). L’interprétation des résultats des données non métriques va dans le sens d’une stabilité de peuplement des ensembles précampaniformes, stabilité qui ne se maintient pas avec l’émergence du Campaniforme. L’hétérogénéité rencontrée lors des périodes tardives est, en général, mise en parallèle avec l’arrivée de nouvelles populations, ou du moins d’un accès modifié au sein de l’aire funéraire (Desideri et Eades 2002, 2004). Ceci va à l’encontre des résultats de l’anthropométrie soutenant une continuité de peuplement jusqu’au Campaniforme. Enfin, une étude basée sur les analyses chimiques des ossements humains a été réalisée par M. Chiaradia, chercheur au Département de minéralogie en Sciences de la Terre à l’Université de Genève (Chiaradia et al. 2003). Il a effectué des analyses des taux isotopiques du plomb et du strontium sur les dents d’individus datant du Néolithique moyen au Bonze ancien provenant de la ville de Sion ainsi que les dents d’un certain nombre de sujets actuels de la région genevoise. Cet auteur a ainsi effectué des analyses des compositions isotopiques du plomb et du strontium tant sur la dentine que sur l’émail de chaque dent, car l’objectif principal de sa recherche était l’étude de la contamination des ossements postmortem et non une étude sur l’histoire du peuplement. Il a tout de même pu mettre en évidence d’après l’émail analysé que tous les individus ont assimilé le même plomb in vivo et qu’ils sont nés et ont probablement vécu toute leur vie au même endroit, sauf un individu, qui aurait migré, pendant sa vie, d’une région géologique différente de celle du Valais. Il s’agit d’un individu campaniforme provenant du dolmen M XI. L’auteur propose également un lieu d’origine qui se situerait en Europe centrale d’après les concentrations en plomb et en strontium de l’émail dentaire du sujet. Les données issues de l’anthropologie semblent s’accorder lorsqu’elles s’intéressent à la situation du début du Néolithique suisse. En revanche, dès le Campaniforme, la situation devient complexe avec des données supportant tantôt une continuité de peuplement, tantôt des différences plus marquées qui pourraient traduire un renouvellement de la population. 67 Chapitre 2 2.6.4. Le Campaniforme en Suisse : les échantillons étudiés Pour la Suisse, les populations analysées se composent d’environ 520 individus répartis sur 8 sites situés chronologiquement entre le Néolithique moyen et le Bronze ancien (figure 13). Ceux-ci sont essentiellement localisés dans la partie occidentale de la Suisse. Seul le site d’Aesch (n° 4) est situé en Suisse septentrionale. Les effectifs sont très variables - de 10 à 120 individus. Sur l’ensemble des sites, les échantillons se répartissent de la manière suivante : - Le Néolithique moyen I est représenté par les sites de Barmaz I (n°1) et II (n°2), Chemin des Collines (n°5) et Avenue Ritz (n°8). - Le Néolithique moyen II est composé des sites de Chamblandes (n°3) et de Corseaux (n°6). - Le Néolithique récent est présent uniquement dans l’aire mégalithique du Petit-Chasseur (n°7) au sein des dolmens MVI (dans sa première phase d’occupation) et MXII. - Le site d’Aesch (n°4) ainsi que deux dolmens de la nécropole du Petit-Chasseur (n°7) (le MVI par réoccupation de la chambre funéraire du Néolithique final et le MXI par édification de leur propre monument) sont à rattacher au Campaniforme. - Le Bronze ancien est constitué de deux échantillons provenant, pour le premier, du site de Barmaz I (n°1) et, pour le second, de plusieurs sépultures de la nécropole du Petit-Chasseur (n°7). Figure 13 : La Suisse, répartition des sites étudiés. Nous avons attribué à chaque site un numéro reporté sur la carte, puis figurent le nom des sites, leur(s) attribution(s) culturelle(s) et, entre parenthèses, leur effectif. 1. Barmaz I : Néolithique moyen (51) ; Bronze ancien (17) / 2. Barmaz II : Néolithique moyen (22) / 3. Chamblandes : Néolithique moyen (74) / 4. Aesch : Campaniforme (environ 50) / 5. Chemin des Collines : Néolithique moyen (24) / 6. Corseaux : Néolithique moyen (44) / 7. Petit-Chasseur : Néolithique récent (dolmen MVI - environ 40 -, dolmen MXII - environ 120 -) ; Campaniforme (dolmen MVI - 10 -, dolmen MXI - 13 -) ; mélanges Néolithique récent - Campaniforme : fosse à incinération (26) ; Bronze ancien (15) / 8. Avenue Ritz : Néolithique moyen (14). 68 Du groupe culturel à l’individu 2.7. L’essentiel en bref Ce chapitre a abordé la présentation aussi bien des régions que des échantillons intégrés dans ce travail. Nous avons tenté de réunir ici une ample portion de la variabilité du Campaniforme, même si elle est de loin non représentative de toute la diversité dont fait preuve ce complexe. Nous avons choisi cinq régions représentant cinq portraits différents de l’occupation campaniforme pendant le 3e millénaire avant notre ère. • Le domaine oriental : la Bohême Cette région envisagée à maintes reprises comme un acteur des plus actifs dans l’histoire du peuplement campaniforme représente l’occupation orientale type. Le Campaniforme se développe lors de la seconde moitié du 3e millénaire avant notre ère, dans une phase finale de l’Enéolithique, en position intermédiaire entre la culture Cordé et le Bronze ancien. Les données provenant de l’archéologie mettent en avant les grandes affinités que le Campaniforme entretient avec les deux cultures qui soit le précède soit lui succède, tant au niveau du rituel funéraire qu’au niveau des vestiges eux-mêmes. L’étude des vestiges humains, en revanche, perçoit une certaine mobilité des individus qui diffère selon l’approche employée. Nous avons réuni ici un échantillon composé de plus de 500 individus et 64 sites provenant des parties septentrionale et centrale de la Bohême. Ils appartiennent à la culture Cordé, Campaniforme et Unetice (Bronze ancien). • Le domaine oriental : la Hongrie L’occupation campaniforme hongroise - le Campaniforme-Csepel - est seule dans l’aire orientale hors substrat cordé. Elle s’intègre au sein d’une phase moyenne du Bronze ancien (BA II) - seconde moitié du 3e millénaire avant notre ère - et se limite à la région entourant Budapest. Différentes interprétations sont avancées pour expliquer l’intrusion limitée du Campaniforme en Hongrie. Pour certains, la genèse de ce complexe résulterait de la migration de petits groupes d’individus provenant de Tchéquie ; pour d’autres, il s’agirait plutôt d’un développement interne de la société du Bronze ancien. Les rares données anthropologiques plaideraient en faveur d’un renouvellement de la population sans exclure des contacts avec les populations contemporaines. Nous avons rassemblé un corpus composé d’une centaine d’individus et de 15 sites localisés dans les parties centro-septentrionale et méridionale de la Hongrie. Les ensembles appartiennent au Campaniforme-Csepel et aux cultures qui lui sont soit contemporaines (Obedà-Pitvaros, Nyirseg et de Somogyvar-Vinkovci), soit successives (Nagyrev, Perjamos). • Le domaine méridional : le sud de la France La fin du Néolithique, marquée par l’émergence d’une multitude de groupes culturels distincts et limités dans l’espace, est subdivisée en deux phases - le Néolithique final et le Chalcolithique. Le Campaniforme s’implante, non de manière uniforme sur l’aire considérée, lors de la dernière phase et coexiste avec les cultures locales dans certains cas. Il se développe pendant la première moitié du 3e millénaire avant notre ère. Les données archéologiques relèvent généralement deux phases d’occupation campaniforme. La première marque une certaine rupture avec les traditions locales, la seconde est une période d’intégration et de cohabitation. Les contextes funéraires compliqués dans cette région n’ont probablement pas induit une multitude d’études anthropologiques. Les données éparses ne permettent pas de proposer une interprétation en terme d’histoire du peuplement. L’échantillon réuni est constitué de près de 700 individus et de 5 sites localisés en grande partie dans la partie orientale de la région. Les ensembles se situent chronologiquement entre le Néolithique final/Chalcolithique et le Bronze ancien. 69 Chapitre 2 • Le domaine méridional : le nord de l’Espagne La péninsule Ibérique est une région importante dans la compréhension du phénomène Campaniforme. La Meseta est un territoire riche en découvertes. Le Campaniforme s’y développe lors de la seconde moitié du 3e millénaire avant notre ère, dans une phase récente du Chalcolithique et coexiste avec lui jusqu’à l’émergence du Bronze ancien. Les données archéologiques vont dans le sens d’une émergence du Campaniforme liée à des transformations de la société de la fin du Néolithique par l’échange de biens socialement valorisés. D’un point de vue des études anthropologiques, aucun élément ne semble défendre un renouvellement ou déplacement de populations avec la genèse du Campaniforme, cependant les études ne les recherchent pas et concernent, avant toute chose, les effets de l’environnement sur les individus. Nous avons constitué un échantillon de près de 180 individus et 14 sites situés dans la partie septentrionale de la Meseta. Les ensembles étudiés sont à rattacher au Néolithique final, au Chalcolithique, au Campaniforme et à l’âge du Bronze. • A l’articulation entre les deux domaines : la Suisse Le territoire helvétique se situe en position intermédiaire entre les domaines méridional et oriental. Les vestiges, inégalement répartis sur l’ensemble de la Suisse, laissent apparaître la dualité des traditions des différentes aires culturelles est-ouest. Le Campaniforme apparaît lors de la seconde moitié du 3e millénaire, à la fin du Néolithique, et laissera place à l’émergence du Bronze ancien. D’après les données archéologiques, la genèse du Campaniforme en territoire helvétique pourrait être le fruit d’influences plus ou moins marquées provenant à la fois de l’aire méridionale - sous forme d’une idéologie - et de l’aire orientale - par migration de populations. Les différentes études anthropologiques constatent une forte homogénéité des populations précampaniformes. En revanche, les résultats pour les populations successives sont plus mitigés, tantôt plaidant en faveur d’une continuité de peuplement, tantôt invoquant un renouvellement de la population. Les populations se composent de plus de 520 individus et de 8 sites provenant essentiellement de la partie occidentale de la Suisse. Cette dernière présente une séquence sans hiatus majeur entre le 5e et le 3e millénaire, c’est pourquoi nous avons sélectionné des échantillons se situant chronologiquement entre le Néolithique moyen et le Bronze ancien. Nous le voyons bien ici, les données sont peu comparables d’une région à l’autre. Ses multiples facettes ne peuvent pas induire une explication unique dans sa genèse, les données anthropologiques illustrent ainsi la complexification du Campaniforme. 70 Chapitre 3 La terminologie dentaire L’objectif de ce chapitre est de dresser un lexique des termes spécifiques à notre recherche. Nous avons délibérément choisi de l’articuler sous forme de texte, et non sous forme de lexique proprement dit, pour en faciliter la lecture. Pour certains, cette partie peut paraître superflue, élémentaire ou encore rébarbative. Nous invitons alors ceux-ci à continuer leur lecture directement au chapitre 4. Pour d’autres, elle ne le sera pas obligatoirement du début à la fin. Nous tenons à avoir de la cohérence dans nos propos, c’est pourquoi certains éléments de base sont répétés ici. 3.1. L’anatomie de la dent La dent (figure 14) est formée de deux parties distinctes, la couronne et la racine. L’extrémité inférieure de la racine se nomme l’apex. Le terme de coronal s’applique à la partie la plus haute de la dent, donc à la couronne et celui d’apical s’applique à la partie la plus inférieure de la dent, donc à la pointe de la racine (l’apex). Figure 14 : Anatomie de la dent 1. couronne 2. racine 3. dentine 4. émail 5. cément 6. cavité pulpaire 7. pulpe 8. collet ou ligne cervicale 9. apex 10. gencive 11. alvéole D’après S. Cugini 1983, p. 24, illustration modifiée. La dent est constituée de plusieurs tissus : La dentine, habituellement appelée ivoire, est le tissu formant le volume de la dent. Ce tissu est dur, élastique, blanc jaunâtre et avasculaire. Il est constitué d’environ 70% d’éléments minéralisés que l’on appelle hydroxyapatite crystallite, de 18% de matière organique, formée principalement de collagène, et de 12% d’eau. Les cellules responsables de la formation de la dentine sont les odontoblastes qui permettent à la dentine de se régénérer (About et al. 2000, 2002). L’émail est le tissu de protection recouvrant la dentine au niveau de la couronne. Il s’agit du tissu le plus minéralisé du corps humain. Il est constitué de plus de 95% d’éléments minéralisés (hydroxyapatite crystallite), de 4% de matière organique et d’1% d’eau. Les cellules responsables de la formation de l’émail sont les améloblastes (About et al. 2000). Ces cellules recouvrent l’émail avant l’éruption de la dent, en revanche, à la fin du processus de croissance elles disparaissent. Ainsi, l’émail ne peut pas se régénérer ou être remplacé (Woelfel et Scheid 1997). 71 Chapitre 3 Le cément est le tissu de protection recouvrant la dentine au niveau de la racine. Il s’agit d’ un tissu dur. Il est constitué d’environ 65% d’éléments minéralisés (hydroxyapatite crystallite), d’une matrice organique, largement composée de collagène, qui s’élève à 23% et de 12% d’eau (Woelfel et Scheid 1997). Les cellules responsables de la formation du cément sont les cémentoblastes (Ruch 2001). La cavité pulpaire se situe à l’intérieur de la dentine et contient le tissu cellulaire, plus connu sous le terme de pulpe, les vaisseaux sanguins et les nerfs (Palosaari 2003). On fait la distinction entre la dentine et la pulpe même si embryologiquement ou encore fonctionnellement ces deux éléments sont similaires (Heiss et al. 1998). L’élément qui les différencie est leur consistance. En effet, l’un est un tissu mou (la pulpe), l’autre un tissu dur (la dentine). Il existe divers termes concernant les zones de contact entre les différents tissus. La jonction cémentémail est la ligne de séparation entre l’émail de la couronne et le cément de la racine que l’on nomme aussi ligne cervicale ou encore collet. Les zones de contact interne entre la dentine et les tissus de protection se nomment respectivement la jonction dentine-émail et cément-dentine (Woelfel et Scheid 1997). Les dents, après éruption, sont fixées chacune dans un processus alvéolaire ou alvéole. La gencive, portion de la muqueuse buccale, recouvre l’arcade alvéolaire et immobilise les dents en les bloquant par leur collet. 3.2. Les dentitions La dentition humaine est diphyodonte indiquant ainsi qu’elle développe deux générations de dents (figure 15). La première est la dentition de lait, déciduale, lactéale ou encore temporaire qui est déjà à moitié formée lors de la naissance et qui se développe pendant les deux premières années de la vie. Celle-ci est remplacée graduellement par la dentition permanente ou définitive, dont la première dent débute son développement peu avant la naissance et la dernière dent est finalement en place vers 20 ans (Hillson 1996). La dentition humaine est également hétérodonte ce qui signifie qu’elle développe des classes dentaires, que l’on nomme indifféremment district ou encore champs, et qui sont localisées selon leur spécialisation (Doro Garetto et al. 1991). Les incisives se situent dans la partie médiane des arcades dentaires. Elles sont facilement reconnaissables car elles ont une forme de spatule - ou encore de lames. Les incisives ont une morphologie compressée et acérée qui servent surtout à inciser et à couper les aliments. Les canines sont des dents à couronne conique et pointue. Cette morphologie leur sert notamment à déchirer et à arracher les aliments. Les prémolaires sont des dents de transition entre les canines et les molaires, les premières sont fréquemment caniniformes et les secondes molariformes. Leur fonction est à la fois de couper et broyer les aliments. Enfin, les molaires situées dans la partie postérieure des arcades ont une morphologie plutôt massive et carrée. Elles servent à broyer et à mâcher les aliments. La dentition est composée de deux arcades dentaires : l’os supérieur - le maxillaire - et l’os inférieur - la mandibule. La moitié d’une arcade se nomme un quadrant. Les quadrants gauches et droits sont séparés par la ligne centrale du corps que l’on nomme le plan sagittal médian. A l’intérieur de chaque quadrant de la dentition déciduale prennent place deux incisives, une canine et deux molaires, qui sont en réalité des prémolaires déciduales. Ce problème provient des noms traditionnels de la dentisterie adoptés sans référence à la paléontologie. Chez les mammifères, la dentition déciduale est constituée d’incisives, de canines et de prémolaires. Selon les espèces, il peut y avoir jusqu’à quatre prémolaires déciduales. Dans le cas de la dentition humaine, les deux molaires déciduales 72 La terminologie dentaire sont équivalentes aux troisième et quatrième prémolaires des autres mammifères 10 (Hillson 1986). Ainsi, la dentition déciduale comprend au total 20 dents. Figure 15 : La dentition humaine A gauche, schéma représentant la dentition permanente et, à droite, schéma de la dentition déciduale. Les différentes classes dentaires sont notées : 1. incisives 2. canine 3. prémolaires 4. molaires Les formules dentaires pour chaque dentition sont placées entre les arcades supérieure et inférieure. D’après S. Cugini 1983, p. 27 et p. 28, illustration modifiée. Pour la dentition permanente, un quadrant se compose de deux incisives, une canine, deux prémolaires et trois molaires. Il y a deux types de dent permanente : celle qui remplace une dent déciduale et que l’on nomme dent successive et celle qui ne remplace aucune dent lactéale, soit les molaires permanentes, que l’on nomme dent additionnelle. Il existe un problème de terminologie similaire pour les prémolaires permanentes qui sont décrites comme les première et deuxième prémolaires et qui sont en réalité des troisième et quatrième prémolaires (Hillson 1986). La dentition définitive comprend au total 32 dents (nombre, qui varie selon les personnes car un individu sans dents de sagesse - correspondant aux troisièmes molaires - aura 28 dents). Les antimères sont les dents symétriques sur une même arcade que l’on nomme également les dents controlatérales. Ainsi, les incisives centrales supérieures gauche et droite sont des antimères. Le terme d’isomère se réfère en revanche à une même dent sur des arcades opposées. Les premières prémolaires supérieure et inférieure sont donc des isomères. 3.2.1. Les termes de positionnement L’arcade dentaire est de forme parabolique, c’est pourquoi les termes d’antérieur et de postérieur ne sont pas réellement appropriés pour décrire l’avant et l’arrière d’une dent. En général, les termes de mésial pour l’avant et de distal pour l’arrière sont utilisés. En ce qui concerne les côtés ou les faces : celle qui se situe du côté de la langue se nomme face linguale, celle du côté des lèvres est la face buccale, vestibulaire ou encore labiale. Les termes d’antérieur et postérieur sont réservés aux dents elles-mêmes : antérieures pour les incisives et les canines, postérieures pour les prémolaires et les molaires. Selon le type de dents auquel nous avons affaire, la surface de mastication se nommera incisale (pour les dents antérieures) et occlusale (pour les dents postérieures). Ce second terme peut tout de même 10 Nous avons adopté les termes traditionnels de la dentisterie dans notre travail afin d’éviter toute confusion. 73 Chapitre 3 s’appliquer à l’ensemble des dents. Certains auteurs, comme E. Marseiller (1984), utilisent également le terme de surface fonctionnelle pour nommer la face de mastication. Les termes d’approximal ou d’interproximal sont utilisés pour définir les zones de contact (mésiale ou distale) entre les dents. Figure 16 : Les termes de positionnement 9 1. face mésiale (vers l’avant de l’arcade) 2. face distale (vers l’arrière de l’arcade) 3. dents antérieures 4. dents postérieures 5. face linguale 6. face buccale, vestibulaire ou labiale 7. face incisale 8. face occlusale 9. face interproximale ou approximale 10. plan sagittal médian D’après S. Cugini 1983, p. 140, illustration modifiée. 3.2.2. Les termes descriptifs morphologiques Les dents montrent un certain nombre de structures, tant positives que négatives, sur l’ensemble de la surface de la couronne. Les surfaces de mastication présentent des petites cavités, appelées également foramen, et des sillons qui prennent leur nom d’après leur localisation. Il existe deux types de sillons. D’une part, les principaux ou fondamentaux qui sont profonds et dissocient les cuspides, et, d’autre part, les secondaires ou accessoires situés sur les cuspides elles-mêmes. La fosse est une dépression sur la surface linguale des dents antérieures et sur la surface occlusale des dents postérieures. Elle se situe en général entre deux sillons. On trouve également sur la couronne diverses structures comme des petits nodules arrondis accessoires - que l’on nomme tubercules - ou encore de crêtes qui prennent leur nom d’après leur position. Toutes les dents possèdent au moins deux crêtes sur les bords proximaux (surface occlusale) que l’on nomme les crêtes marginales qui, selon leur position, seront distale ou mésiale. Il existe toute une série de termes spécifiques à l’anatomie de la racine. La furcation est la zone de division des racines chez une dent pluriradiculaire (possédant donc plusieurs racines). Ainsi, cette zone se déclinera en bifurcation pour une dent biradiculaire (deux racines) et en trifurcation pour une dent triradiculaire (trois racines). Les divisions créées par le développement de sillons qui peuvent parfois fractionner une racine se nomment les radicules. Une radice est une petite racine accessoire. 3.2.3. Les termes spécifiquement réservés aux dents antérieures Des termes sont spécifiquement réservés pour décrire les différentes parties des dents antérieures. Chaque partie des dents antérieures se subdivise en trois zones distinctes. La couronne, en vue longitudinale, se divise en trois parties. La zone cervicale correspond à la région proche de la jonction cément-émail. La zone incisale se situe au sommet de la dent. La partie intermédiaire se nomme la zone médiane. En largeur, les trois zones se nomment respectivement zones distale, médiane et mésiale. La racine se subdivise en zones apicale (extrémité de la racine), médiane et cervicale. 74 La terminologie dentaire Le cingulum est un élargissement ou une protubérance se situant sur la partie cervicale de la surface linguale de la couronne. Le mamelon correspond au tubercule sur le bord incisal des incisives. En général, trois tubercules y sont présents. 3.2.4. Les termes spécifiquement réservés aux dents postérieures Il existe des termes particuliers pour la description des différentes parties des dents postérieures. Comme pour les dents antérieures, les diverses fractions des dents postérieures sont subdivisées en trois zones précises. En vue longitudinale, la couronne se divise en zones cervicale, médiane et occlusale. En ce qui concerne les termes de subdivisions de la largeur de la couronne et de la racine, les trois régions correspondent à celles décrites pour les dents antérieures (c.f. chapitre 3.2.3.) Les cuspides sont des proéminences sur la surface des molaires et des prémolaires. Le sulcus est une dépression au centre de la surface occlusale. Les différentes cuspides des molaires peuvent être nommées de diverses manières selon le système de terminologie adopté. Il existe trois systèmes qui sont, en général, employés indifféremment par les auteurs. Les tableaux ci-dessous (figures 17 et 18) mettent en correspondance les trois systèmes afin d’avoir une vision globale des termes adoptés pour la description des cuspides des molaires. Termes de positionnement Système tribosphénique Système anthropologique Cuspide mésiolinguale Cuspide mésiovestibulaire Cuspide distovestibulaire Cuspide distolinguale -- Protocone Paracone Métacone Hypocone Métaconule Cuspide 1 Cuspide 2 Cuspide 3 Cuspide 4 Cuspide 5 Figure 17 : Présentation des différents systèmes employés pour la description des molaires supérieures. Termes de positionnement Système tribosphénique Système anthropologique Cuspide mésiovestibulaire Cuspide mésiolinguale Cuspide centrovestibulaire Cuspide distolinguale Cuspide distovestibulaire -Cuspide centrolinguale Protoconide Métaconide Hypoconide Entoconide Hypoconulide Entoconulide Métaconulide Cuspide 1 Cuspide 2 Cuspide 3 Cuspide 4 Cuspide 5 Cuspide 6 Cuspide 7 Figure 18 : Présentation des différents systèmes employés pour la description des molaires inférieures. Le premier système utilise les termes de positionnement décrits ci-dessus, il s’agit de la notation la plus précise dans la dénomination des cuspides. Le deuxième correspond au système tribosphénique de H.F. Osborn 11. Les auteurs utilisent assez fréquemment ce système pour nommer des traits dentaires correspondant à la présence d’une cuspide. Finalement, nous pouvons utiliser la terminologie employée par l’anthropologie dentaire. Les cuspides sont tout simplement numérotées, l’avantage de ce système est celui d’être très facile, mais il a l’inconvénient d’être peu précis. 11 Ce système est utilisé en paléontologie pour déterminer les cuspides présentes chez les mammifères. 75 Chapitre 3 3.3. Les systèmes de notation dentaire Différents systèmes de notation dentaire sont utilisés dans la littérature. Il est important de les connaître dans la mesure où souvent les dents sont codées et donc leur compréhension est peu accessible. Les plus fréquents sont présentés en figure 19. Système universel dentition permanente maxillaire dentition déciduale mandibule maxillaire mandibule troisième molaire deuxième molaire première molaire deuxième prémolaire première prémolaire canine incisive latérale incisive centrale incisive centrale incisive latérale canine première prémolaire deuxième prémolaire première molaire deuxième molaire troisième molaire deuxième molaire première molaire canine incisive latérale incisive centrale incisive centrale incisive latérale canine première molaire deuxième molaire droite 1 2 3 4 5 6 7 8 25 26 27 28 29 30 31 32 A B C D E P Q R S T gauche 16 15 14 13 12 11 10 9 24 23 22 21 20 19 18 17 J I H G F O N M L K Système de Palmer ou Zsigmondy droite 8 7 6 5 4 3 2 1 1 2 3 4 5 6 7 8 E D C B A A B C D E gauche 8 7 6 5 4 3 2 1 1 2 3 4 5 6 7 8 E D C B A A B C D E Système international droite 18 17 16 15 14 13 12 11 41 42 43 44 45 46 47 48 55 54 53 52 51 81 82 83 84 85 Figure 19 : Présentation des différents systèmes de notation dentaire. 76 • Le système universel (Universal Numbering System), mis en place par Parreidt en 1882, a été adopté par l’association américaine dentaire en 1975 (Woelfel et Scheid 1997). Le principe est relativement simple : pour les dents permanentes un chiffre est attribué allant de 1 à 32. La numérotation débute sur la troisième molaire supérieure droite, la troisième molaire supérieure gauche correspond à 16, la troisième inférieure gauche à 17 et la droite à 32. Pour la numérotation de la dentition déciduale, des lettres de l’alphabet sont utilisées allant de A à T. Elle débute par la deuxième molaire supérieure droite selon le même principe. • Les systèmes de Palmer (1861) et de Zsigmondy (1870) complètent la numérotation des dents par des parenthèses spécifiant le quadrant auquel elles appartiennent. Le symbole signifie supérieur droit, inférieur gauche, etc. Les dents permanentes sont notées de 1 à 8. La notation débute par l’incisive centrale et se termine par la troisième molaire pour chaque quadrant. Pour les dents déciduales, une lettre de l’alphabet est attribuée allant de A à E selon le même principe. gauche 28 27 26 25 24 23 22 21 31 32 33 34 35 36 37 38 65 64 63 62 61 71 72 73 74 75 La terminologie dentaire • Le système international ou F.D.I. (Fédération Dentaire Internationale) attribue à chaque dent deux chiffres. Le premier correspond à la fois à la dentition, à l’arcade et au côté. Ainsi, la dizaine signifie dentition permanente-supérieure-droite et la septantaine dentition déciduale-inférieure-gauche. Le second chiffre correspond à la dent ellemême. La numérotation débute par l’incisive centrale et se termine par la troisième molaire pour chaque quadrant. Les dents permanentes sont notées de 1 à 8 et les dents déciduales de 1 à 5. Dans notre travail, nous n’avons choisi aucun des systèmes décrits ci-dessus. Nous avons, en effet, préféré une terminologie simple et facilement compréhensible de notation dentaire, souvent utilisée par l’école américaine. Pour la dentition permanente, chaque dent est codée de la manière suivante : • Une lettre de l’alphabet en majuscule correspond à la première lettre de leur nom (M= molaire, P = prémolaire, C = canine et I = incisive). • Un chiffre est ajouté pour spécifier la dent de la classe dentaire à laquelle nous avons affaire. Il est placé en exposant ou en indice, ce qui nous permet également d’indiquer l’arcade à laquelle la dent appartient (M1 = première molaire supérieure, P2 = deuxième prémolaire inférieure...). • Enfin, la latéralisation de la dent est spécifiée par les lettres g (gauche) ou d (droite). Exemple : M3 g soit la troisième molaire inférieure gauche. Pour la dentition déciduale le même système a été adopté : • Une lettre de l’alphabet en minuscule correspond à la première lettre de leur nom (m = molaire, c = canine et i = incisive). • Un chiffre est ajouté pour spécifier la dent de la classe dentaire à laquelle nous avons affaire. Il est placé en exposant ou en indice, ce qui nous permet également d’indiquer l’arcade à laquelle la dent appartient (m1 = première molaire supérieure, i2 = deuxième incisive inférieure...). • Enfin, la latéralisation de la dent est spécifiée par les lettres g (gauche) ou d (droite). Exemple : m2d soit la deuxième molaire supérieure droite. Cette notation sera celle adoptée dans la plupart des cas ici (texte et annexes). Néanmoins, selon les logiciels utilisés, cette terminologie peut parfois être modifiée. De ce fait, il est possible de trouver dans les tables de ce travail des noms de variables en majuscule ou en minuscule indépendamment de la dentition considérée. Exemple : T1M3D ou t1m3d, signifiant tous deux le même trait observé sur la même dent, soit le trait 1(absence congénitale) observé sur la troisième molaire droite. Il s’agit tout simplement d’un problème lié au logiciel utilisé et à la version de ce dernier que nous ne pouvons malheureusement pas contrôler. Ainsi, pour distinguer les deux dentitions, nous avons simplement ajouté un d (qui peut être en minuscule ou en majuscule) avant l’initiale de la dent pour la dentition déciduale. Exemple : T28DI2G ou t28di2g, soit le trait 28 (extension de l’émail de la surface vestibulaire) observé sur l’incisive latérale déciduale gauche. De même, les prémolaires dans certaines tables peuvent être codées par l’initiale P ou PM. 77 Chapitre 3 3.4. Quelques définitions supplémentaires La dentition humaine peut parfois présenter des dents surnuméraires, une rétention de dents déciduales, une transposition dentaire, qui est l’inversion de la localisation de deux dents ; ou encore l’absence de dents (absence congénitale), qui peut se traduire sous deux formes : l’anodontie qui implique l’absence de l’entière dentition, ou l’hypodontie qui concerne uniquement quelques dents. La synodontie se réfère à une fusion totale de deux dents ; lorsqu’elle n’est que partielle (n’impliquant pas la couronne) le terme de schizodontie est employé. L’hypercémentose est un excédent de cément au niveau de la racine. L’hypoplasie est une perturbation affectant l’émail, provoquée par des troubles répétés pendant l’enfance (nutritionnels ou infectieux). Cette lésion se manifeste par la présence de stries sur la couronne des dents. Cette atteinte altère l’émail dentaire et expose plus fortement ces dents à la carie. Les dents isolées, ainsi que les maxillaires et les mandibules disjoints sont, généralement, regroupés sous le terme de vracs. Nous n’irons pas plus loin dans ce rappel des termes utiles pour la compréhension de ce travail. Ainsi, nous ne décrirons pas chaque dent car tel n’est pas le propos ici. Cependant de très belles et complètes synthèses, d’où nous avons tiré en grande partie ces informations, peuvent être consultées à cette intention. Nous citerons, parmi d’autres ouvrages, S. Cugini (1983), G.C. van Beek (1983), E. Marseiller (1984), S. Hillson (1996), T. Doro Garetto et al. (1991), M.M. Ash (1993), J.B. Woelfel et R.C. Scheid (1997), L. Scheuer et S. Black (2000) et T.D. White et P.A. Folkens (2005). 78 Chapitre 4 Les traits non métriques dentaires : des indicateurs des distances entre les populations Les traits non métriques dentaires s’observent sur l’ensemble de la dentition. L’un des avantages de ces caractères est de pouvoir être observés sur le vivant. Cet élément n’est pas négligeable car il permet d’acquérir une meilleure connaissance de leur valeur biologique - situation relativement rare en anthropologie physique. De plus, la dent est l’une des parties les plus résistantes du squelette et représente souvent l’élément le mieux conservé en milieu archéologique. Ces deux points permettent de penser que l’étude des traits non métriques dentaires peut se révéler une approche des plus intéressantes lors de recherches touchant les populations du passé. L’intérêt pour l’observation des variantes morphologiques dentaires n’est pas récent puisque celles-ci étaient déjà décrites par les anatomistes grecs (Ossenberg 1976). De ce fait, il semble utile de débuter ce chapitre par un bref historique des recherches. Il n’est en aucun cas exhaustif et relate principalement l’état de la recherche quant à la mise en place des différentes définitions des variables et leur application pour la caractérisation des populations. Ensuite, nous montrerons que la morphologie dentaire s’avère être un outil attractif dans les analyses concernant la compréhension des relations entre populations. Pour déterminer si les traits non métriques dentaires sont de bons estimateurs des distances biologiques, nous aborderons différents aspects. D’abord, nous nous attacherons à la valeur biologique de ces variables. Il faut avant tout pouvoir montrer que le développement de la dentition est contrôlé par les gènes. Ensuite nous nous intéresserons à leur mode de transmission. Puis, nous verrons quels sont les effets de l’environnement sur leur développement. Et finalement, nous tenterons de mesurer le rôle des gènes dans leur expression. Puis, nous nous intéresserons à la fiabilité du système d’observation, l’élément important le plus discuté. Enfin, nous vérifierons si ces variables, appliquées sur des populations actuelles, s’avèrent suffisamment sensibles pour être considérées comme de bons indicateurs des distances biologiques. 4.1. Un historique des recherches L’étude des variantes morphologiques dentaires débute au 19e siècle par leur description dans les différentes populations d’alors. Au tournant de ce siècle-là, les principes fondamentaux de l’ontogénie dentaire se mettent en place. En 1920, A. Hrdlicka est le premier à classer le degré d’expression d’un trait morphologique dentaire, les crêtes linguales marginales (variable plus connue sous le terme d’incisives en pelle) (figure 20), en sériant ces variations au sein de plusieurs populations humaines. Par la suite, il étend son travail en étudiant d’autres traits dentaires (Hrdlicka 1921). Cette époque est marquée par l’émergence de travaux portant sur les variations morphologiques dentaires en vue d’études comparatives. C.J. Shaw, en 1927, décrit la morphologie dentaire des populations bantoues et San. M.S. Goldstein, en 1931, s’intéresse à la morphologie des molaires chez les Inuits et encore C.T. Nelson, en 1938, décrit les variations dentaires des Indiens Peco. P.M. Butler, en 1939, met en place son concept des gradients rencontrés dans la dentition comme celui de champs morphogénétiques. Les travaux de A.A. Dahlberg (1945) concernent les variations des traits dentaires d’Indiens du sud-ouest de l’Amérique. En 1949, P.O. Pedersen étudie la dentition des populations arctiques. 79 Chapitre 4 Figure 20 : Incisive latérale supérieure gauche présentant un développement de crêtes marginales sur les bords de sa surface linguale. Figure 21 : Première molaire supérieure gauche montrant sur sa face mésiolinguale un tubercule accessoire plus connu sous le terme de tubercule de Carabelli. Les années cinquante marquent un pas décisif pour l’anthropologie dentaire. Les recherches se concentrent sur la génétique et la morphogénèse des variations morphologiques de la couronne. B.S. Kraus (1951) explique le mode de transmission du tubercule de Carabelli (figure 21). H. Grüneberg (1952) est le premier à affirmer que les variantes anatomiques sont des variables quasi-continues. A partir des années soixante, l’anthropologie dentaire devient une discipline à part entière. K. Hanihara, en 1961, met au point des plaques de référence pour les variantes morphologiques des dents déciduales en se basant sur les travaux de A.A. Dahlberg. En 1963, D.R. Brothwell publie un ouvrage sur les différents aspects de l’anthropologie dentaire. En 1965, le premier symposium international sur la morphologie dentaire a lieu au Danemark, sous l’égide de A.A. Dahlberg, P.O. Pedersen et V. Alexandersen réunissant des chercheurs de différents horizons, notamment des anthropologues, des généticiens, des dentistes et des paléontologues. Cette rencontre donne naissance, d’une part, à d’autres symposiums qui ont eu lieu tous les trois ans, et d’autre part, à une grande quantité d’articles sur la morphologie dentaire. Dans les années soixante-dix, N.E. Morton crée des méthodes statistiques testant les systèmes polygéniques (Morton et al. 1971). C.G. Turner développe, en 1970, des gradations standardisées pour le développement de certains traits de la couronne des molaires. Ce travail aboutit à la création du système d’enregistrement des traits dentaires ASU (Arizona State University). Des études sur la transmission des caractères dentaires (Christian et al. 1974, Berry 1978…) s’intensifient en se basant sur l’étude de jumeaux et de sujets apparentés. En 1978, J.H. Osborn propose un nouveau modèle pour expliquer le développement des classes dentaires et leurs gradients, qu’il nomme modèle du Clone. Dès lors, les études sur les variantes dentaires se multiplient et suivent, en général, trois directions : • L’établissement de nouveaux critères de classification (Alvesalo et al. 1975, Sciulli 1977, Aas et Risnes 1979a/b, Turner et al. 1991…). • La caractérisation des populations actuelles, notamment des études concernant l’Océanie (Smith et al. 1982, Townsend et Brown 1980, Townsend et al. 1990…), l’Amérique (DeVoto et Perrotto 1971, Sofaer et al. 1972b, Escobar et al. 1976, Brewer-Carias et al. 1976, Scott et al. 1983…), l’Afrique (Chagula 1960, Greene 1982, Haeussler et al. 1989…) ou encore l’Asie (Manabe et al. 1992, Higa et al. 2003…) laissant l’Europe faire figure de parent pauvre avec quelques rares études (Brabant 1967, Berry 1976…). • La compréhension des aspects biologiques des variantes dentaires comme les mécanismes de l’odontogenèse, leur mode de transmission ou encore les influences environnementales sur leur développement (Harris et Bailit 1980, Townsend et Brown 1980, Keene 1982, Saunders et Mayhall 1982, Townsend et al. 1986, Nichol 1989…). L’emploi des traits non métriques dentaires en contexte archéologique est relativement récent. Les études concernent principalement la caractérisation, les mouvements de populations et l’histoire du peuplement ancien (Greene 1982, Lukacs 1983a/b, Lukacs et Walimbe 1984, Irish et Turner 1990, Coppa et al. 1998, Irish 1997, Desideri 2001, 2005…). Ces variations anatomiques sont incorporées dans les études internes depuis une quinzaine d’années seulement (Sjøvold 1977, Alt et Vach 1992, Alt 1997, Eades 1997, 2003, Piera 2000…). 80 Les traits non métriques dentaires 4.2. Les traits non métriques dentaires Lorsque l’on parle des traits non métriques dentaires, on considère toute variation anatomique observée sur l’ensemble de la dentition permanente et déciduale. On rencontre dans la littérature une série de termes qui sont employés indifféremment comme trait, caractère, variante, variable, anomalie ou encore figure, auxquels on ajoute un adjectif pour les distinguer des traits métriques tel qu’épigénétique, discret, non métrique, morphologique, discontinu ou encore à seuil. On se réfère, en général, à des caractères qui peuvent être présent ou absent, ou qui manifestent différents stades de développement. Ils peuvent prendre la forme de structures positives comme la présence de tubercules ou de crêtes accessoires, ou négatives tels des sillons. Ils se manifestent également sous la forme de variations dans le nombre, la position ou encore la taille des cuspides et des racines (figure 22). Figure 22 : Quelques exemples de traits non métriques dentaires observés sur la dentition permanente : (A) nombre de cuspides des molaires inférieures, en haut 4 cuspides et en bas 5 / (B) à droite, incisive latérale présentant deux sillons traversants, l’un médian et l’autre distal ; à gauche incisive latérale exprimant un tubercule dentaire sur sa surface linguale / (C) une extension de l’émail est présente sur la molaire supérieure / (D) molaire supérieure en vue occlusale exprimant un tubercule de Carabelli / (E) nombre de racines d’une première prémolaire supérieure, à gauche monoradiculaire, et à droite biradiculaire / (F) radix paramoralis présente sur une première molaire supérieure. La terminologie des traits dentaires est variable et diffère selon les auteurs. Prenons, par exemple, les différents synonymes que l’on peut rencontrer dans la littérature pour deux de ces traits: • Pour le tubercule de Carabelli, les termes équivalents sont tubercule mésiolingual, protostyle, anomalie de Carabelli ou encore tuberculus anomalous. • Pour l’entoconulide, les termes équivalents sont cuspide 6, tuberculum sextum ou encore tuberculum accessorium posteriore internum. Le flou dans la dénomination des traits a souvent induit une confusion entre la définition des variables et les termes employés, et a parfois conduit à des erreurs d’observation. Il existe une centaine de traits dentaires dont un peu moins d’une quarantaine ont fait l’objet d’études plus poussées touchant les aspects purement biologiques de leur développement, leur mode de transmission ou encore leur héritabilité. 4.2.1. L’ontogénie dentaire Le développement de la dentition déciduale débute entre la 6e et 7e semaine de la vie prénatale. Après une phase de croissance simultanée des dentitions temporaires et permanentes, l’odontogenèse se termine entre 18 et 25 ans par la rhizagenèse (formation des racines) et l’éruption des 3e molaires (Ruch 2001). Des travaux in vivo et in vitro en laboratoire sur des tissus embryonnaires dentaires ont permis d’avoir une meilleure compréhension des mécanismes et des processus de l’odontogenèse (Wang 2004). Les études concernant le développement dentaire des rongeurs, en particulier des souris, ont largement contribué aux connaissances que l’on peut avoir aujourd’hui de l’organogenèse en général. 81 Chapitre 4 4.2.1.1. Le développement dentaire L’ontogénie dentaire débute pendant les stades initiaux du développement cranio-facial et peut être subdivisée en trois phases : l’initiation, la morphogenèse et la différenciation (ou histogenèse) (figure 23). Il s’agit d’un processus complexe impliquant des interactions coordonnées - séquentielles et réciproques entre l’épithélium et le mésenchyme (Pispa et Thesleff 2003). Les différents tissus ont ainsi deux origines embryologiques. L’émail est d’origine épithéliale et les autres tissus (dentine, cément, pulpe et ligaments alvéolodentaires) dérivent du mésenchyme (Wheater et al. 2001). Les étapes initiales de la croissance de la dentition sont similaires à celles des autres organes dérivés de l’ectoderme comme les cheveux ou encore les glandes mammaires (Tummers 2004). Le développement coronaire est généralement subdivisé en trois phases : bud, cap et bell stages. Cette classification ne traduit pas des modifications fonctionnelles significatives mais décrit l’évolution de la morphologie (Ten Cate 1998). Dans des endroits spécifiques des arcs mandibulaire et maxillaire, l’épithélium oral donne naissance aux lames dentaires (dental lamina) et le mésenchyme se condense. Les lames engendrent par la suite des épaississements épithéliaux localisés envahissant le mésenchyme que l’on appelle les germes dentaires (dental placode) et qui correspondent à la localisation des futures dents. Ces épaississements vont produire les bourgeons dentaires (bud stage). Par division des cellules épithéliales et mésenchymateuses, les bourgeons croissent et se transforment en capuchon dentaire (cap stage). La forme concave de ce dernier entoure le mésenchyme, la future pulpe dentaire. Les capuchons se convertissent graduellement par histogenèse en organe de l’émail. On assiste également à l’apparition d’une structure provisoire qui est un arrangement de cellules aptotiques (autodestruction programmée) et de cellules de remplacement regroupées sous le terme de nœud de l’émail (enamel knot). Nous verrons plus loin le rôle fondamental de ce groupe de cellules. Une amplification des processus de croissance et d’histogenèse permettent aux capuchons de se transformer en cloche dentaire (bell stage). La forme des dents est déterminée par la combinaison de prolifération des cellules et par mouvements cellulaires. Pendant le stade en cloche, la différenciation des cellules (odontoblastes et améloblastes) donne naissance aux divers tissus constituant la dent (Mitsiadis 2001). La différenciation des odontoblastes débute au sommet des cuspides et évolue en direction apicale. Ces cellules synthétisent et sécrètent les constituants organiques de la dentine12 (Ruch 2001). La différenciation des améloblastes débute, comme pour les ondotoblastes, au sommet de la surface occlusale. Elles synthétisent et produisent, quant à elles, les constituants de l’émail 13. La rhizagenèse prend place à la fin du développement coronaire. L’activité des odontoblastes se poursuit en produisant la partie organique de la dentine. Cette expansion vers le bas des cellules se nomme la gaine de Hertwig. Elle déterminera la taille finale de la racine. En contact avec la gaine, les cémentoblastes débutent leur différenciation et produisent le cément qui va graduellement la remplacer (Stevens et Lowe 1996). 4.2.1.2. Les aspects moléculaires Les analyses concernant les aspects génétiques de la dentition étaient jusque dans les années quatre-vingtdix limitées à quelques mesures de l’héritabilité (Polly 2006). Ces dernières années, les recherches ont nettement amélioré la compréhension du développement moléculaire de la dent. Ainsi, on peut affirmer aujourd’hui que la formation de la dentition est sous le contrôle d’un certain nombre de gènes qui agissent à divers endroits et qui interviennent à différents moments de l’ondontogenèse déterminant la position et la forme de chaque dent (Thesleff et Nieminen 1996). Plus de 300 gènes sont impliqués dans le 12 Les principaux constituants organiques de la dentine sont les collagènes, les protéoglycanes, les glycoprotéinessialoprotéines, les phosphoprotéines, les protéines riches en acide-carboxyglutamique, l’améline, les métalloprotéases, les lipides, les facteurs de croissance et les protéines sériques (Ruch 2001). 13 Les principaux constituants organiques de l’émail sont les amélogénines, les énamélines, les améloblastines, la tufteline, les sialophosphoprotéines, la laminine-5, les enzymes protéolytiques, la phosphatase alcaline, les lipides et les protéines sériques (Ruch 2001). 82 Les traits non métriques dentaires développement de la dentition comme le montrent, entre autres, les différents travaux issus du programme de Biologie développementale de l’institut de Biotechnologie de l’Université d’Helsinki concernant le développement dentaire et craniofacial coordonné par I. Thesleff. Les travaux ont surtout concerné la reconnaissance des gènes qui sont impliqués dans la formation de la dentition et leur interaction (Jernvall et Thesleff 2000, Thesleff 2003...). Figure 23 : Le développement de la dent. En haut, synthèse des différentes étapes de la croissance dentaire, de l’initiation à l’éruption. En bas, synthèse des divers facteurs connus actuellement intervenant lors des différents stades de développement de la dent. D’après I. Thesleff 2003, p. 1647-1648. Les interactions entre les différents tissus cellulaires responsables de l’odontogenèse sont sous l’influence de divers agents composés de facteurs de transcription, de facteurs de croissance et de molécules de la matrice extracellulaire (figure 23). Parmi les facteurs de transcription, on trouve certaines familles de gènes comme les homéogènes, les oncogènes ou encore des gènes liés au développement embryonnaire. Les homéogènes 14 ou les gènes homéotiques définissent d’une part la position et les spécificités de chaque organe, et de l’autre ils contrôlent la synthèse des facteurs de transcription qui codifient l’activité des gènes de structure. Les principaux homéogènes impliqués dans le développement de la dentition sont les Alx, les Barx, les Dlx, les Lhx, les Msx ou encore les Pitx. Les proto-oncogènes15 sont des gènes intervenant dans la régulation des mitoses. Au sein des gènes liés à l’embryogenèse, on peut citer les familles Pax, Lef-1 ou encore Eda (Mitsiadis 2001, Cobourne et Sharpe 2003). Les facteurs de croissance, qui sont des molécules de signalisation, interviennent non seulement dans la division mais également dans la migration et la différenciation cellulaire. Les principaux facteurs de croissance sont la super-famille des TGFβs comprenant les TGFβs, les BMPs, les activines, la follistatine, la famille des EGF comprenant le polypeptide EGF et le TGFα, la famille des FGF qui sont des facteurs de croissance 14 Un gène homéotique est un gène régulateur qui commande la destinée des groupes de cellules au cours du développement embryonnaire (Campbell et Reece 2004). 15 Les proto-oncogènes sont des gènes impliqués dans le contrôle de la division cellulaire. Un dérèglement de leur expression par mutation provoque une tumeur (multiplication excessive de cellules), on les appelle alors des oncogènes. 83 Chapitre 4 fibroclastique, la famille des neurotrophines, les HGF, les PDGF, les molécules de midkine et de pléiotrophine ou encore les acides rétinoïques (Thesleff et Nieminen 1996, Mitsiadis 2001). Enfin, les molécules de la matrice extracellulaire participent à la division, la migration, l’adhésion des cellules et à l’histogenèse (Mitsiadis 2001). Les loci des gènes jouant un rôle dans l’odontogenèse sont peu à peu localisés sur les différents chromosomes (Sperber 2004). C’est lors d’expériences impliquant la mutation ou la suppression d’un gène entraînant des malformations, voire une absence totale ou partielle des dents, que l’on peut mettre en avant son implication dans le développement (Nieminen et al. 1995, Pirinen et al. 1996, 2001, Axelson et al. 2003, Axenovich et al. 2003, Miletich et Sharpe 2003, Pemberton et al. 2006...). Le rôle précis de chaque gène connu impliqué dans l’odontogenèse fait l’objet d’études qui sont encore à leur début. Néanmoins certains travaux ont permis de cerner certaines fonctions de ces molécules (Qju et al. 1997, Ferguson et al. 1998, Line 2001). Par exemple, l’homéogène Barx1 induit les bourgeons dentaires à la molarisation (Tucker et al. 1998), les homéogènes Msx1 et Alx3 déterminent la morphologie incisiforme des dents et la région de chevauchement entre les homéogènes Msx1 et Dlx interviennent dans les districts des canines et des prémolaires (Sperber 2004). Plusieurs travaux ont montré qu’en amplifiant l’expression de l’ectodysplasine (gène Eda), le nombre de cuspides augmente, leur forme et leur position se modifient et le sujet présente également des dents surnuméraires (Kangas et al. 2004, Tucker et al. 2004). P.T. Sharpe et son équipe ont émis l’idée de l’existence d’un code homéoboîte16 odontogénique (odontogenic homeobox code) impliqué dans la détermination de la localisation et du type de dent (Sharpe 1995, 2001, Tucker et Sharpe 1998, Tucker et al. 2004). Ce concept a été médité à partir d’expériences ayant démontré le rôle des gènes homéotiques Hox dans la position des vertèbres le long de l’axe du corps de l’embryon (Horan et al. 1995)17. Le nœud de l’émail (enamel knot) a un rôle important et serait même un centre de signalisation et d’organisation qui gérerait les informations concernant la morphogenèse et la croissance des cuspides (Vaahtokari et al. 1996). Le nœud d’émail primaire se développe à la transition entre le stade du bourgeon et du capuchon réglant la progression de la morphogenèse et situé à la base de la première cuspide qui se développera (Harris et Dinh 2006). Les nœuds secondaires apparaissent à l’étape suivante (stade de la cloche) au niveau de la future localisation des autres cuspides (Thesleff 2003). Il semblerait qu’ils codifient la forme et la taille des cuspides (Tucker et Sharpe 1998, Cobourne et Sharpe 2003). Y. Kassai et al. (2005) ont découvert que l’intensité de l’expression de l’ectodine, un inhibiteur des protéines morphogénétiques osseuses (BPMs) du noeud de l’émail, a un effet non seulement sur ce dernier mais également sur la forme des cuspides et le nombre de dents. Aujourd’hui, des modèles informatisés mêlant activité des gènes, interactions développementales et variations phénotypiques ont été conçus dans une perspective de compréhension de la formation des cuspides et de la variation morphologique dentaire (Salazar Ciudad et Jernvall 2002). Though the exact functions of genes in morphogenesis still remain unknown, somes genes [...] have closer spatial correlation to morphology than others, and it is possible that such genes are more relevant to the patterning and evolution of the species-specific shapes than other genes [...] which are otherwise essential for the normal tooth development (Keränen et al. 1998, p. 485). Nous sommes loin d’avoir élucidé tous les mécanismes du développement dentaire, toutefois les études se multiplient et les acquis sont déjà importants. Maintenant savoir quels sont les gènes impliqués dans l’expression ou le stade d’un trait non métrique dentaire, la voie est ouverte même si le chemin à parcourir est encore long avant de pouvoir y répondre. 16 Une homéoboîte (homeobox) est une section d'ADN située dans les gènes assurant le contrôle du développement. Elle est constituée de 180 paires de bases codant pour un domaine protéique, l'homéodomaine. Les gènes à homéoboîte sont des régulateurs de transcription (activateurs ou inhibiteurs) contrôlant une cascade d'autres gènes indispensables à la morphogenèse d’un organe précis (Mitsiadis 2001). 17 Les gènes à homéoboîte Hox sont positionnés les uns après les autres le long de certains chromosomes. L’ordre de leur arrangement représente la position des différents organes qu’ils contrôlent. Ces gènes sont, par l'intermédiaire des protéines régulatrices qu’ils produisent, des gestionnaires de l’organisation anatomique (Mitsiadis 2001). 84 Les traits non métriques dentaires 4.2.1.3. Les aspects biologiques Les mammifères possèdent tous un même ensemble de dents qui se différencie selon le type d’alimentation. Prenons la dentition des carnivores, elle est spécialisée puisque leur nourriture nécessite principalement des dents acérées. Les rongeurs, quant à eux, possèdent une paire d’incisives à croissance continue et trois paires de molaires. Au contraire, l’homme, qui a une alimentation omnivore, possède une dentition non spécialisée. Elle est définie de diphyodonte et d’hétérodonte. Toutefois, on trouve des gradients de la dentition où chaque dent ressemble à sa voisine. Il est courant de trouver dans la dentition humaine des canines inférieures incisiformes, ou encore, des prémolaires caniniformes ou molariformes, montrant ainsi un continuum morphologique entre les différentes classes dentaires (Scott et Turner 1997). On utilise le terme de gradient de l’expression pour expliquer les variations morphologiques que l’on rencontre dans les classes dentaires. Les chercheurs expliquent ces variations en terme de champs et de clones. Ces concepts sont utiles pour comprendre les variations de la dentition, mais les mécanismes impliqués ne sont pas véritablement compris. Le concept de champs morphogénétiques dentaires dérive de l’analyse de P.M. Butler (1939) sur la morphodifférenciation des dents des mammifères fossiles du Cénozoïque. Il met au point le concept de gradients de la dentition comme celui de champs morphogénétiques (figure 24). Ce chercheur postule que les dentitions des mammifères peuvent être différenciées le long de l’axe antéro-postérieur en trois champs morphogénétiques équipotents qui correspondent aux groupes des incisives, des canines et des molaires. Chaque groupe est sujet à un champ d’influence génétique distinct qui gouverne la taille et la forme de la dent dans le groupe (Butler 1978). De plus, l’intensité de l’influence morphogénétique est meilleure sur une dent particulière dans chaque groupe, et cette dent est la plus stable dans sa taille et sa forme. Les dents les plus mésiales de chaque classe morphologique auront une plus grande concentration en substance et seront celles qui sont soumises au plus fort contrôle génétique. Au contraire, les dernières dents qui se développent dans un groupe (ou district), sont celles qui montrent les plus grandes composantes environnementales. A.A. Dalhberg, en 1945, reprend le concept des champs morphogénétiques mis en place par P.M. Butler, en y intégrant un district supplémentaire, celui des prémolaires. Figure 24 : Schéma représentant le modèle des champs morphogénétiques - Field theory - mis en place par P.M. Butler (1939). D’après R. Ten Cate 1998, p. 91. J.H. Osborn, en 1978, développe le modèle du Clone pour expliquer le développement des classes dentaires et des gradients d’expression (figure 25). La situation de départ est identique à celle du modèle des gradients morphogénétiques (théorie des champs). La région spécialisée qui se développe se nomme ici clone ou primordium (une population de cellules). Toutes les dents d’un district dentaire dérivent d’une même région cellulaire qui se développe à partir de cellules souches (stem precursor), et qui produit séquentiellement les dents dont la configuration dépend du potentiel de forme des cellules d’origine. Chaque clone ou primordium développe une structure qui sera ensuite dupliquée selon le nombre de dents par district. La première dent développée, qui est en général la dent en position mésiale, exprimera toutes les instructions initiales du primordium. En revanche, les suivantes ne les exprimeront que partiellement par perte d’informations. 85 Chapitre 4 Figure 25 : Schéma reproduisant le modèle du Clone - Clone theory -, proposé par J.H. Osborn (1978), concernant le développement des dents postérieures (les molaires). D’après R. Ten Cate 1998, p. 91. La théorie du Champ morphogénétique présuppose l'uniformité initiale des ébauches dentaires et l'apparition graduelle de dents de forme et de taille différentes en fonction d’interactions avec des concentrations en gradient d’un morphogène hypothétique. En revanche, la théorie du Clone implique que chaque ébauche dentaire initiale est intrinsèquement différente et sa morphogenèse contrôlée de manière indépendante (Ruch 2001). Pour P.M. Butler (2001), les deux théories sont partiellement justes. De nombreux auteurs ont souligné le fait que ces modèles décrivent, mais n’expliquent pas, les régularités dans un système qui présente des gradients (Scott et Turner 1988, 1997, Ruch 2001, Tyrrell 2001...). Les progrès croissants des études concernant les bases moléculaires de la dentition pourraient à long terme permettre de favoriser l’une ou l’autre de ces théories, de les combiner, voire de les infirmer. Mais, en attendant, les concepts des théories du Champ et du Clone sont utiles pour comprendre les variations morphologiques que l’on rencontre dans la dentition. 4.2.2. Le mode de transmission des traits non métriques dentaires Plusieurs travaux se sont intéressés au mode de transmission des traits dentaires. Pour affirmer que la fréquence phénotypique est conforme à la fréquence génotypique, il faut démontrer que ces caractères suivent un mode de transmission simple par l’action de gènes dominants, récessifs ou codominants (modèle de Mendel). Au départ, les chercheurs ont tenté de valider ce modèle pour tous les traits dentaires, même pour ceux qui présentent des expressions variables. B.S. Kraus (1951) est le premier à déterminer le mode de transmission du tubercule de Carabelli par l’étude de sujets apparentés. Il en conclut que l’expression de cette variable résulte de l’action de gènes codominants, en admettant tout de même que d’autres facteurs génétiques doivent avoir une certaine influence sur sa transmission. Différents auteurs ont essayé tant bien que mal de démontrer que les différents traits étudiés suivent le modèle mendélien, mais ils ont rencontré de grandes difficultés pour valider ce système comme seul mode de transmission des traits dentaires (Turner 1967, 1969, DeVoto et Perrotto 1971, Escobar et al. 1976…). En 1960, J.H. Edwards pensait que les mécanismes impliqués dans la transmission de traits qui s’expriment uniquement par l’absence et la présence sont bien plus complexes. H. Grüneberg (1952) a été le premier à affirmer que certains phénotypes discontinus, refusant de suivre le modèle mendélien, obéissent à des modes de transmission polygénique. Il a nommé ces variables quasi-continues. Le concept de variables quasi-continues a été introduit en anthropologie par R.J. Berry et A.C. Berry (1967) et A.C. Berry (1976) pour justifier l’emploi de variables non métriques dans les études entre populations. Ainsi, différents chercheurs, notamment D.H. Goose et G.T.R. Lee (1971), ont commencé à 86 Les traits non métriques dentaires accepter des modèles de transmission polygénique, selon lesquels un certain nombre de gènes interagissent, chacun avec un effet additif, pour permettre au trait de s’exprimer. Les traits non métriques dentaires sont considérés comme des variables quasi-continues par opposition à celles dites discontinues caractérisées par un mode de transmission simple, et à celles dites continues qui possèdent des mécanismes complexes d’hérédité (Grüneberg 1952). Le modèle de la variation quasicontinue développé par D.S. Falconer (1965) permet de comprendre la relation entre le phénotype et le génotype des variables quasi-continues. Ce modèle (figure 26) présente une distribution des traits qualitatifs morphologiques au sein d’une population. La distribution dérive de la prédisposition de chaque individu à exprimer un trait, combinée à des facteurs environnementaux qui rendent plus ou moins possible cette manifestation (Tyrell 2000). Tous ces facteurs suivent une courbe normale à l’intérieur d’une même population. Le seuil (threshold), qui est le point après lequel tous les individus exprimeront le trait, varie selon les populations; et la susceptibilité (liability) est la position relative d’un individu sur la distribution en relation avec le seuil. Figure 26 : Adaptation du modèle des variables à seuil de D.S. Falconer (1965) pour un trait non métrique hypothétique. D’après A.J. Tyrell 2000, p. 291. Des méthodes statistiques, les analyses complexes de ségrégation (complex segregation complex), ont été développées pour évaluer le mode de transmission d’un caractère au sein d’une ascendance (Morton et al. 1971, Morton et MacLean 1974, Elston et Stewart 1971). Plusieurs anthropologues ont appliqué ces méthodes aux traits dentaires. D. Kolakowski et al. (1980) ont été les premiers à utiliser les analyses complexes de la ségrégation pour évaluer le mode de transmission du trait de Carabelli. Ils ont observé ce trait sur 358 familles nucléaires des îles Salomon. La variance du trait de Carabelli serait pour la plupart due à l’effet d’un gène majeur. Néanmoins ces auteurs estiment qu’une petite part de la variance serait le résultat d’action de gènes additifs situés sur différents loci. C.R. Nichol (1989, 1990) a également effectué ce type d’analyse mais sur un nombre plus important de traits dentaires (24 dont 17 variables non métriques appartenant au système ASU) par l’étude de moulages appartenant à 83 familles nucléaires d’Indiens Pima. Sur l’ensemble des traits étudiés, 14 d’entre eux possèderaient un gène majeur responsable de la transmission des traits. Ces gènes majeurs seraient soit dominants, soit récessifs - respectivement dans 10 et 3 cas -, ainsi qu’un indéterminé18. Quatre autre traits 18 Les traits sous l’influence d’un gène majeur dominant sont la protubérance, l’insetting et le sillon traversant, les crêtes linguales marginales, les crêtes vestibulaires marginales, le trait de Carabelli, les variations des cuspides linguales, l’hypoconulide, le métaconulide et le torus maxillaire. La forme de l’arcade maxillaire est sous le contrôle d’un gène majeur récessif. La dominance du gène majeur des cuspides accessoires n’a pu être définie. 87 Chapitre 4 pourraient également être, sans grande certitude, sous l’influence d’un gène majeur19. Les six derniers sembleraient bien être contrôlés par plusieurs gènes 20. Certains traits non métriques sont recensés dans le catalogue de V.A. McKusick (1990) concernant les phénotypes à hérédité mendélienne. On trouve dans cet ouvrage plus d’une quarantaine de variables dentaires notamment des anomalies liées au développement de la dentine et de l’émail, telle la dysplasie ou encore la dentinogenis imperfecta, la synodontie, les dents surnuméraires ou l’éruption dentaire avant la naissance. En ce qui concerne les variables observées dans notre étude, six traits sont inventoriés dans le catalogue et posséderaient tous un mode de transmission autosomal dominant. Il s’agit de l’absence congénitale (divers types d’hypodontie), du degré de rotation des incisives centrales supérieures, des crêtes linguales marginales des incisives supérieures, du sillon traversant (dens invaginatus), de l’odontome, et du trait de Carabelli. Même s’ils sont listés au sein de l’ouvrage, le mode de transmission de ces différentes variables n’est pas établi de manière très sûre. Cependant, les résultats des différentes études montrent que le mode de transmission des caractères non métriques dentaires semble bien être de type multifactoriel, cependant il est bien moins complexe que celui présent pour les variables continues. Certains traits semblent être sous l’influence d’un gène majeur, d’autres, en revanche, seraient sous le contrôle de gènes agissant chacun avec un effet additif à divers endroits et à différents moments du développement de la dent. 4.2.3. Les effets de l’environnement Aujourd’hui, il n’est plus à démontrer que le développement de la dentition est contrôlé par l’action des gènes, cependant on suppose que l’environnement doit avoir une certaine influence sur la morphologie dentaire. D’une part, des expériences in vivo et in vitro, toujours sur des rongeurs, ont permis de mieux cerner les éléments ou les facteurs environnementaux agissant sur le développement de la dentition. D’autre part, des études portant sur des populations humaines migrantes ou sur des ensembles comprenant diverses générations ont fourni des indications intéressantes. Ainsi, des expériences en laboratoire ont établi que le régime alimentaire de la génitrice enceinte ou qui allaite peut avoir une certaine influence sur la taille des dents de sa progéniture. A.G. Searle (1954a/b) a démontré d’après des expériences sur des souris qu’une progéniture dont la mère enceinte a suivi un régime alimentaire pauvre en protéines aura des troisièmes molaires de dimensions réduites. Le travail de R.S. Bader (1965) a lui aussi indiqué que le milieu maternel joue un rôle essentiel dans la détermination de la taille des dents, voire de leur absence congénitale. En effet, il a mis en évidence qu’une déficience en vitamine A réduit la taille de la dentition de façon significative. Pour R.M. Grainger et al. (1959), le type d’alimentation pendant le développement dentaire influence la taille de la dentition. B.J. Krueger (1962, 1966) a testé l’influence du boron, du fluor et du molybdène sur la morphologie dentaire en formation des rongeurs. Le premier élément ne semble pas avoir d’effet sur le développement de la dentition, les autres substances produisent des traumatismes du calcium au niveau de la matrice de l’émail. Toutefois les effets se limitent souvent à des atteintes post-natales et sont moindres in utero (Tenzcar et Bader 1966, Larson et Bader 1976). Les études concernant les facteurs environnementaux pouvant avoir un effet sur la taille de la dentition humaine ne sont pas nombreuses. Elles ont également permis de confirmer que la taille des dents peut être influencée par l’environnement. Par exemple, I.J. Møller (1967) a réalisé une synthèse des travaux concernant les effets des éléments traces sur la morphologie dentaire. Il en ressort avant tout que la quantité de fluor dans l’eau potable joue 19 Les traits sous l’influence incertaine d’un gène majeur seraient la convexité labiale des incisives, le tubercule accessoire distal (qu’il nomme métaconule) des molaires supérieures, l’entoconulide et le protostylide des molaires inférieures. 20 Les traits sous l’effet additifs de plusieurs gènes (transmission de type polygénique) seraient le degré de rotation des incisives, la crête accessoire distale des canines, le dessin du sillon des molaires inférieures, le pli mésiolingual de la première molaire inférieure, la forme de l’arcade mandibulaire ainsi que le torus mandibulaire. 88 Les traits non métriques dentaires un rôle certain sur la dentition. Une teneur élevée en fluor confère une taille supérieure à la dentition combinée à des cuspides plus arrondies et des sillons moins profonds. H.L. Bailit et B. Sung (1968) ont tenté de déterminer si le milieu maternel influence le développement de la dentition permanente sur un ensemble comprenant une centaine de familles vivant à Boston. Ils ont constaté que le poids à la naissance du sujet ou encore l’âge de la mère peuvent avoir des effets sur la taille de la dentition. De même, S.M. Garn et al. (1980) ont testé plusieurs facteurs, des conditions maternelles (diabète, hypertension et hyperthyroïdie) et des conditions développementales (durée de gestation, poids et taille à la naissance), qui pourraient avoir des effets systématiques sur les dimensions des dents déciduales et permanentes. Leurs résultats montrent que chacun des facteurs analysés influencent la taille des dents. Par exemple, le diabète de la mère a pour conséquence une taille dentaire inférieure à la moyenne de sa progéniture, inversement l’hypertension ou l’hyperthyroïdie confère à la dentition une taille supérieure. De plus, les effets se retrouvent aussi bien sur la dentition déciduale que sur la permanente. D. Kolakowski et E.F. Harris (1981) ont souligné l’existence d’effets différentiels de facteurs environnementaux sur la taille des dents antérieures selon le diamètre considéré. C.F. Ebeling et al. (1973) ont travaillé sur les changements séculaires de la taille des dents parmi des populations suédoises. Ils ont remarqué une augmentation de la taille à travers le temps qui est probablement due, selon les auteurs, à des interactions génético-environnementales. Tous ces travaux ont montré que la taille des dents, dont le taux de plasticité est comparable à celui de la stature, peut être influencée par divers facteurs environnementaux. Il reste tout de même une grande interrogation concernant le rôle de l’environnement dans l’expression des traits non métriques dentaires. Une seule étude s’est penchée sur l’effet d’un élément sur l’expression d’un trait dentaire. Il s’agit du travail de G.J. Cox et al. (1961) sur les effets liés à l’ingestion de fluor, pendant le développement dentaire, sur la taille du tubercule de Carabelli. Ils n’observent aucune influence directe du fluor sur l’expression de cette variable. Certains chercheurs se sont intéressés à d’autres éléments qui pourraient éventuellement influencer le développement des traits dentaires. Il semble, selon G.R. Scott et C.G. Turner (1997), que la moyenne annuelle de la température, l’humidité, les radiations ultraviolettes ou encore l’altitude n’aient aucun impact sur le développement de ces caractères. Il se trouve que la majorité de ces effets agissent aussi in utero mais que les mammifères soient très bien protégés contre ce type d’atteintes pré-natales (Tyrell 2000). G.W. Lasker (1945) a étudié la morphologie dentaire de populations chinoises et de leurs descendants immigrés aux Etats-Unis. Les incidences populationnelles des traits dentaires observés dans les deux ensembles étudiés sont tout à fait similaires. La migration pendant le Moyen Age de populations scandinaves provenant d’un climat maritime modéré dans des régions climatiques bien différentes telles le Groenland et l’Islande fournissent des indications des plus intéressantes quant aux modifications de la morphologie. En effet, il semblerait que les variations du crâne, notamment les torus palatins et mandibulaires, bien que fortement héréditaires, subissent des contraintes environnementales significatives (Halffman et al. 1992). En revanche leur morphologie dentaire est conforme à celle de leurs ascendants (Scott et Alexandersen 1992). De même, G.R. Scott et C.G. Turner (1997) constatent que les morphologies dentaires des descendants des colonisateurs européens sont identiques à celles de leurs ancêtres et demeurent très différentes de celles des populations indigènes. Actuellement, le problème est de savoir quels sont les éléments qui ont un effet sur les traits dentaires et surtout quelles en sont leurs conséquences. On sait que les traits dentaires ne sont probablement pas totalement protégés des influences environnementales, néanmoins leur effet semble mineur et leur impact n’est pas significatif sur les fréquences populationnelles (Scott et Turner 1997). Pour G.R. Scott et R.H.Y. Potter (1984), une adaptation phénotypique du système dentaire produirait des mutations dans les fréquences génotypiques au sein d’une population qui migrerait dans une région impliquant des modifications environnementales (changements alimentaires par exemple). Pour ce faire, ces ajustements phénotypiques doivent comporter des avantages sélectifs. Ainsi, l’incidence des traits dentaires dériveraient d’échanges génétiques plutôt que de fluctuations environnementales (Eades 2003). 89 Chapitre 4 4.2.4. L’héritabilité des traits non métriques dentaires Le phénotype est l’ensemble des caractéristiques observables chez un individu. Il est déterminé en partie par les gènes et en partie par l’environnement. La part du déterminisme génétique dans l’expression des traits dentaires a suscité un grand intérêt chez les anthropologues, cependant ils ont souvent mal interprété ce que le calcul de l’héritabilité pouvait, en réalité, apporter comme résultat. En effet, il ne faut pas confondre l’héritabilité au sens étroit du terme, qui correspond au rapport entre la variance génétique additive et la variance phénotypique (soit la mesure du déterminisme génétique d’un caractère), de l’héritabilité au sens large du terme. Cette dernière correspond à la part de la variabilité causée par les effets génétiques, dans la variabilité totale observée pour un caractère dans une population. Il s’agit d’une proportion dans une population et n'a aucun sens pour un individu. L’héritabilité mesure le degré de transmission de ce caractère d’un parent à ses descendants. Ainsi, son calcul n’est pas un indice du déterminisme génétique, mais exprime la confiance à accorder au phénotype des individus en tant qu’appréciation de leur valeur génétique additive (Falconer 1974, p. 128). Pour les traits dentaires, lorsqu’on parle d’héritabilité, on se réfère généralement à cette définition. Pour calculer l’héritabilité21 d’un caractère, il faut tenir compte de différents éléments. D’une part, la transmission de la variable doit être de type polygénique, c’est-à-dire chaque gène agissant avec un effet additif pour permettre au trait de s’exprimer. De l’autre, l’héritabilité est spécifique à la population et à l’environnement où elle a été calculée (Falconer 1974). La parenté des individus prescrit le degré auquel on s'attend à ce que leur similitude soit déterminée par des facteurs génétiques (Kohn 1991). La ressemblance entre apparentés a été un moyen pour estimer l’héritabilité des traits dentaires. A l’aide du calcul de coefficient de corrélation entre parents et enfants, des analyses ont pu évaluer l’héritabilité de certains traits dentaires (Sofaer et al. 1972a, Scott 1973, Alvesalo et al. 1975, Blanco et Chakraborty 1977, Sharma 1992...). Les deux caractères les plus étudiés ont été le tubercule de Carabelli et les incisives en pelle. Leur héritabilité a été estimée à travers plusieurs études basées sur différentes populations vivant dans des environnement distincts (Scott et Turner 1997). Une héritabilité similaire pour chacun des traits a été constatée dans les différents ensembles étudiés. Selon G.R. Scott et C.G. Turner (1997), les variables non métriques dentaires montrent un haut degré d’héritabilité, se situant entre 0.4 et 0.8. Une estimation du déterminisme génétique peut être effectuée d’après des analyses de concordance des traits dentaires chez les jumeaux. Si l’on part de l’hypothèse que les gènes définissent l’expression de ces variables, on suppose que pour des jumeaux parfaits ou monozygotes, toutes différences d’expression devraient être attribuées à l’environnement. De plus, les concordances entre les expressions des faux jumeaux ou jumeaux dizygotes devraient être moins élevées. Ainsi, des études portant sur les analyses de concordance entre jumeaux (Ludwig 1957, Berry 1978, Kaul et al. 1985, Townsend et al. 1986, 1990, Liu et al. 1998...) ont permis de confirmer cette présomption. A. Lundström (1963) a réalisé un exercice relativement intéressant par l’étude d’un échantillon constitué des moulages de la dentition de plus de 120 paires de jumeaux. Il a tenté de définir d’après la morphologie dentaire la zygosité, soit la détermination de la monozygosité ou de la dizygosité des paires d’individus. Il a réussi à classer correctement 95% des paires de jumeaux étudiées. J.C. Boraas et al. (1988) ont étudiés des jumeaux monozygotes séparés à la naissance, qui ont ainsi vécu dans des environnements complètement différents (communauté, habitudes alimentaires diverses...), présentant des concordances très élevées de l’expression des différents traits dentaires. L’étude de la variance peut également être employée pour évaluer le déterminisme génétique des jumeaux (Scott et Potter 1984, Boraas et al. 1988, Townsend et Martin 1992...). Finalement, les différentes études ont permis de conforter l’idée que la plupart des caractères non métriques dentaires semble disposer d’une grande composante héréditaire. 21 L’héritabilité est égale à deux fois la corrélation dans l’expression d’un trait. Quand la valeur est proche de 1, cela signifie qu’une large proportion de la variance phénotypique d’un caractère au sein d’une population est due à des effets additifs génétiques. Ainsi elle n’indique pas la proportion totale des facteurs génétiques dans l’expression d’un trait car l’influence d’un gène majeur dans l’expression d’un trait, par exemple, ne sera pas comptabilisée (Falconer 1965). 90 Les traits non métriques dentaires 4.3. La fiabilité du système d’enregistrement La fiabilité du système d’observation est un élément fondamental, voire crucial, dans chaque étude, quelle que soit l’approche utilisée. Il faut pouvoir garantir une normalisation des modes d’observation car il s’agit d’apporter une crédibilité aux données qui seront ensuite analysées; sans celle-ci, les résultats peuvent être contestables. On a souvent remis en doute l’objectivité de l’enregistrement des traits non métriques dentaires qui sont avant tout des variables qualitatives et qui peuvent être enregistrées de diverses manières : • La dichotomie (A=0/P=1) : elle se limite à l’alternative présence ou absence d’un trait. • La gradation (A=0/P=1, 2...) : on attribue à la présence d’un trait divers stades de développement. • La mesure (A=0/P=10, 20... mm) : les expressions qualitatives des variables sont transformées en données quantitatives. La dichotomie a été la première approche adoptée par les chercheurs dans les études morphologiques dentaires datant de la fin du 19e siècle et du début du 20e siècle. Certains traits s’observent uniquement par leur absence ou leur présence, mais d’autres présentent différents degrés d’expression. Avec des données dichotomisées pour des variables qui présentent des gradations, les chercheurs sont limités dans leurs moyens d’analyse car ils perdent des informations essentielles comme la variabilité de l’expression des traits morphologiques. Si la dichotomie s’est avérée ne pas être le meilleur moyen d’enregistrement pour ce genre de données, la quantification ne l’est pas non plus, même si, en théorie, elle représente l’approche la moins discutable. En effet, selon G.R. Scott et C.G. Turner (1997), la majorité des variables dentaires n’est pas adaptée à la mesure, car les traits montrent des variations tant dans la taille que dans la forme. Ils présentent parfois des niveaux d’expression si fins qu’ils sont impossibles à quantifier. La meilleure méthode d’enregistrement se limite, à l’heure actuelle, aux différentes gradations qui peuvent être observées 22. Ces échelles contiennent les caractéristiques quantitatives et qualitatives de la morphologie dentaire. A. Hrdlicka, en 1920, a été le premier à montrer l’avantage de la subdivision de l’expression des traits dentaires. Ce type de données prend en compte l’expressivité variable qui peut être observée sur une grande partie de ces caractères. Cependant, certains chercheurs ont insisté sur la subjectivité dans l’évaluation de l’expression des caractères morphologiques dentaires. Ils estiment que la présence d’un trait ne peut se subdiviser car l’utilisation de gradations amplifie les erreurs d’observation (Sofaer et al. 1972a). En effet, pendant longtemps l’absence de standardisation des définitions des variables et l'incapacité d’observer les différentes variations des traits dentaires ont été un réel problème pour les chercheurs. Ils ont été obligés de constater la difficulté de comparer des données provenant de différents observateurs. En 1950, A.A. Dalhberg a allégé ce problème en créant une série de moulages montrant la variabilité de plusieurs traits dentaires. Il a été suivi par K. Hanihara, qui, dans les années soixante, a développé des plaques de référence pour les variantes non métriques des dents déciduales (Hanihara 1961). Si ces moulages ne sont pas disponibles, ils existent sous forme de photographies, publiées en 1963 dans son article « Crown characters of the deciduous dentition of the Japanese-American hybrids ». Par la suite, C.G. Turner, en collaboration avec l’Université de l’Etat de l’Arizona, a créé un ensemble de plaques de référence (figures 27 et 28) regroupant une vingtaine de traits sur les dents permanentes (système ASU) (Turner et al. 1991). L’utilisation des moulages est incontournable, elle réduit en effet les différences d’appréciation entre observateurs ou par un même chercheur à différents moments de la recherche. 22 Pour établir une échelle, les chercheurs se basent sur une détermination de l’expression de la plus faible à la plus forte. Il faut fixer, dès le départ, les expressions minimum et maximum, puis délimiter les expressions intermédiaires. 91 Chapitre 4 Figure 27 : Exemple de plaques de référence du système ASU (Arizona State University) reproduisant les différents stades que l’on peut observer dans l’expression des incisives en pelle. A gauche, moulage présentant les différentes variations rencontrées pour l’incisive centrale supérieure ; à droite, moulage pour l’incisive latérale supérieure. Figure 28 : Exemple de plaques de référence du système ASU (Arizona State University) reproduisant les différents stades que l’on peut observer dans l’expression de certaines cuspides supplémentaires sur les molaires inférieures. A gauche, moulage exposant les variations rencontrées pour l’entoconulide ou 6 e cuspide ; à droite, moulage du métaconulide ou 7e cuspide. G.R. Scott (1975), dans une étude sur les variations dentaires de groupes d’Indiens du sud-ouest de l’Amérique, a mesuré les erreurs intra-observateurs en enregistrant un même corpus à deux reprises. Il s’est intéressé au pourcentage d’erreurs dans la détermination de présence ou d’absence d’un trait et au pourcentage d’erreurs d’attribution des expressions phénotypiques. Il trouve des taux de concordance variant entre 85,7% et 100% avec une moyenne de 95,2%. En ce qui concerne les gradations, il trouve une moyenne de 84,3% pour l’assignation d’un même stade. Ceci nous montre la difficulté d’évaluer la manifestation d’une chose à un endroit précis et de savoir si cette chose est ou n’est pas le trait à observer. C.G. Turner et K. Hanihara (1977) ont étudié 85 moulages de dentitions Ainu en s’intéressant aux problèmes interobservateurs. L’un des points intéressants dans ce travail est la détermination du tubercule de Carabelli, cuspide mésiolinguale des molaires supérieures. Selon les résultats de K. Hanihara, les Ainu se révèlent très similaires aux Asiatiques tandis que les données de C.G. Turner indiquent le contraire, aucune affinité n’étant constatée. Ces différences seraient dues, d’après les auteurs, à la difficulté de l’un des deux chercheurs à estimer correctement les expressions faibles du trait. Cette étude nous montre que les différences entre les observateurs peuvent parfois aboutir à des interprétations complètement contradictoires. C.R. Nichol et C.G. Turner (1986) ont publié l’étude la plus détaillée sur les erreurs intra- et interobservateurs. C.R. Nichol a observé à trois reprises un corpus et C.G. Turner une seule fois. Ils ont enregistré 45 traits dentaires dont 37 de format gradué possèdent des plaques de référence et 8 sont observés uniquement sur leur présence et leur absence. Ils ont à leur disposition 4 séries d’observations qu’ils vont comparer les unes aux autres. Ils recensent huit valeurs leur permettant d’évaluer la concordance d’enregistrement des traits dentaires de leur échantillon. La première valeur consiste à évaluer les différences d’enregistrement d’un trait pendant une session d’observation qui n’est pas enregistré pendant une session différente qu’ils nomment observed only one session pourcentage. Cette situation se retrouve en moyenne à 7% lors d’une comparaison intra-observateur et à 13% entre deux observateurs. Ils calculent ensuite toute une série de pourcentages de discordance d’attribution d’expression : discordance absolue (variant scoring percentage), variation de plus d’un stade de différence (>1 grade variant scoring percentage), les différences moyennes de stades brutes (net mean grade difference) et en valeur absolue (absolute mean grade difference). Ils effectuent finalement des tests de similarité entre les distributions afin d’estimer si les différences rencontrées sont trop importantes pour être aléatoires. Nous verrons plus en détail lors de l’évaluation de notre propre concordance (chapitre 5) que les résultats de l’étude de C.R. Nichol et C.G. Turner (1986) situent les divergences 92 Les traits non métriques dentaires interobservateurs entre 25% et 30% et intra-observateurs entre 10% et 30% pour les différentes valeurs mesurées. Le problème ne se résume donc pas seulement à l’estimation du degré d’expression, mais aussi à l’évaluation de la présence ou non d’un trait. Les plaques de référence ont été mises en place par souci d’objectivité. De plus, elles permettent d’identifier les expressions les plus faibles des traits, qui est le problème le plus souvent soulevé. L’objectif à atteindre, durant la phase d’enregistrement, est une systématisation des modes d’observation afin d’assurer la réplicabilité des résultats. Idéalement, deux séries d’observations effectuées par deux chercheurs différents devraient donner sensiblement les mêmes résultats avec un minimum d’erreurs intra- et interobservateurs. Il semble impossible d’obtenir 100% de concordance dans des échantillons qui sont observés à plusieurs. Les plaques de référence développées par A.A. Dahlberg, K. Hanihara ou encore C.G. Turner permettent tout de même d’attribuer des stades avec plus de précision. Finalement, lors d’une étude, mis à part l’utilisation des plaques de référence et de définitions précises, il est indispensable de pouvoir valider ses données en testant avant tout sa propre réplicabilité et, dans la mesure du possible, sa concordance avec d’autres chercheurs. 4.4. Le potentiel de ces variables dans les études entre populations Le dernier élément à vérifier est de déterminer si les traits non métriques dentaires sont de bons indicateurs des distances biologiques entre les populations. Pour le faire il faut pouvoir appliquer ces variables dans des études portant sur des ensembles actuels et ceci à différents niveaux de différenciation, c’est-à-dire de l’étude locale à celle intercontinentale. Nous n’évoquerons pas ici l’utilisation des traits non métriques dans les études internes ou familiales puisqu’il s’agit d’un niveau d’analyse que nous n’aborderons pas dans ce travail. Différents travaux de synthèse peuvent être consultés comme ceux de K. Alt et ses collaborateurs (Alt et Vach 1991, 1995, Alt 1997, Alt et al. 1995, 1998) ou encore de S. Eades (1997, 2003...). Plusieurs études ont permis de mettre en évidence le potentiel de la morphologie dentaire en tant que discriminant des distances biologiques à l’échelle continentale (Shaw 1927, Goldstein 1931, Nelson 1938, Pedersen 1949, Moorrees 1951…). En effet, les fréquences de certains traits dentaires varient fortement parmi les cinq principales régions géographiques. Pour ne citer que quelques exemples, le tubercule de Carabelli est une variable courante chez les Européens, les incisives en pelle sont très fréquentes chez les Asiatiques, la cinquième cuspide des molaires supérieures et la sixième cuspide des molaires inférieures sont des traits habituels chez les Aborigènes ou encore la septième cuspide que l’on rencontre presque exclusivement chez les populations africaines. Certains auteurs ont même employé la fréquence des traits morphologiques dentaires pour définir des complexes comprenant entre 5 et 10 traits permettant de distinguer les populations à un niveau intercontinental. K. Hanihara a défini en 1967 un complexe asiatique ou Mongoloid dental complex23, d’après les incidences très élevées de certains traits présents sur la dentition déciduale des populations asiatiques et amérindiennes. Ce complexe a par la suite été étendu sur la dentition permanente et finalement subdivisé par C.G. Turner (1987) en deux groupes distincts : les Synodontes (comprenant les populations du nord-est de l’Asie et les populations amérindiennes) et les Sundadontes (caractérisant les populations du sud-est asiatique, de la Polynésie et de la Micronésie)24. J.T. Mayhall et al. (1982) ont, 23 Le complexe se compose des variables suivantes pour la dentition déciduale : les crêtes linguales marginales des incisives supérieures, le métaconule (tubercule accessoire distal) sur les secondes molaires supérieures, le pli mésiolingual, le protostylide et le métaconulide sur les secondes molaires inférieures. Complété ensuite pour la dentition permanente par les crêtes linguales marginales, le pli mésiolingual, le protostylide, l’entoconulide et le métaconulide sur les premières molaires inférieures. 24 Cette dichotomie se base sur les divergences d’incidences de huit traits : les crêtes linguales et vestibulaires des 1e incisives supérieures, la 1e prémolaire monoradiculaire, l’extension de l’émail de la 1e molaire, l’absence congénitale de la 3e molaire, le pli mésiolingual et la racine triradiculaire de la 1e molaire inférieure et le nombre de cuspides de la 2e molaire inférieure. Pour plus d’informations, se référer à l’article de C.G. Turner (1990). 93 Chapitre 4 quant à eux, mis en évidence un complexe caucasoïde ou Caucasoïd dental complex25 comprenant un set de traits dentaires habituellement observés dans les populations d’origine européenne. G.C. Townsend et al. (1990) ont défini un ensemble de variables fréquemment présentes chez les Aborigènes qu’ils ont nommé le complexe australien ou Australian dental complex26. Enfin, J.D. Irish (1995, 1997, 1998) a mis en évidence un set de traits permettant de caractériser les populations sub-sahariennes qu’il a nommé subSaharan African dental complex27 et un ensemble nord africain nommé North African dental complex28. Cependant, si les traits non métriques dentaires sont de bons indicateurs lorsque l’échelle de différenciation est grande, on a souvent remis en doute leur pouvoir discriminant dans le cas contraire. Pour tenter de savoir si les données non métriques dentaires peuvent être un bon indicateur des distances biologiques à une échelle plus fine, les chercheurs les ont comparées à d’autres estimateurs comme la métrique, la linguistique, la répartition géographique ou encore les données génétiques. C.G. Turner (1969) a été l’un des premiers chercheurs à avoir effectué une telle analyse. Il a comparé les fréquences des données dentaires aux fréquences d’estimateurs sanguins d’Indiens d’Amérique du sudouest. Les résultats ont montré que les concordances entre ces deux types de données ne sont pas parfaites, mais elles sont néanmoins largement comparables. J.A. Sofaer et al. (1972b) ont confronté des distances de données dentaires de diverses tribus d’Indiens d’Amérique avec des distances géographiques, des estimateurs des différences génétiques calculées à partir des fréquences des groupes sanguins et les variantes des protéines sériques. Dans un premier temps, les correspondances entre les données dentaires et les différents indicateurs se sont révélées relativement mauvaises. En effet, les auteurs se sont heurtés à différents problèmes méthodologiques, notamment en rencontrant des difficultés dans la réplication de leurs observations, qui est l’un des éléments de base pour la validation des données. Ils ont donc décidé d’éliminer les variables posant de sérieux problèmes de standardisation et ont réalisé une nouvelle analyse. Celle-ci s’est révélée bien plus fructueuse avec de très bonnes corrélations entre les différents estimateurs. L’analyse de C.A. Brewer-Carias et al. (1976) a tenté de déterminer la concordance d’une matrice de distances, calculée d’après des données non métriques dentaires, avec des matrices de distances basées sur des marqueurs génétiques pour évaluer le degré de microdifférenciation au niveau local (entre différents villages d’Indiens Yanomama). De très bonnes correspondances ont été obtenues entre les distances dentaires, les distances génétiques et la proximité géographique des différents ensembles étudiés. Dans le même esprit, l’étude de H.R. Palomino et al. (1977) a tenté de saisir la valeur des données non métriques dentaires à travers l’étude de diverses populations et ceci à divers niveaux de différenciation. Le corpus étudié résulte d’une compilation de données récoltées dans la littérature et les résultats qui en découlent sont mitigés. Il semble que les données dentaires soient de bons discriminants entre populations 25 Le complexe comprend des fréquences élevées du degré de rotation des incisives supérieures et du tubercule de Carabelli sur les molaires supérieures et du protostylide sur les molaires inférieures et de l’absence ou de fréquences relativement basses des crêtes linguales marginales des incisives supérieures, d’odontome sur les prémolaires, d’entoconulide et de métaconulide sur les molaires inférieures. 26 Le complexe a été défini d’après la fréquence très élevée de l’entoconulide sur les molaires inférieures ainsi que par la présence du métaconule (tubercule accessoire distal) et du tubercule de Carabelli sur les molaires supérieures et de fréquences très basses pour le protostylide et le métaconulide sur les molaires inférieures. 27 Le complexe se compose de fréquences élevées de la crête mésiale de la canine supérieure, de la première prémolaire biradiculaire supérieure, du tubercule de Carabelli sur les molaires supérieures, de la racine de Tomes sur la première prémolaire inférieure, le dessin du sillon en Y et la racine biradiculaire des deuxièmes molaires inférieures. Des fréquences très basses non seulement du degré de rotation et de crêtes marginales vestibulaires des incisives centrales supérieures, mais aussi l’extension de l’émail des premières et de l’absence congéniatale des troisièmes molaires supérieures. 28 La morphologie dentaire de ce complexe est très proche de celle européenne, toutes deux sont simples et n’expriment que rarement des expressions très élevées des traits dentaires, à l’exception de ceux qui les caractérisent. Le complexe nord-africain est donc défini par des fréquences très basses du degré de rotation des incisives supérieures, du métaconule (tubercule accessoire distal) et du pli de la cuspide mésiolinguale de la première molaire supérieure, de la racine biradiculaire de la première prémolaire et celle triradiculaire de la deuxième molaire supérieures, la racine de Tomes de la première prémolaire inférieure, le dessin du sillon en Y et la racine biradiculaire des deuxièmes molaires inférieures. Ce complexe est également caractérisé par des fréquences élevées du sillon traversant des secondes incisives supérieures et de l’absence congénitale des troisièmes molaires supérieures. 94 Les traits non métriques dentaires lorsque l’échelle de différenciation est grande. En revanche, dans le cas contraire, elles semblent bien moins efficaces. Cela étant dit, G.R. Scott et A.A. Dahlberg (1982) estiment qu’il faut considérer comme contestable les résultats de cette étude, l’analyse ayant été effectuée avec des données publiées ce qui rend impossible le contrôle de la standardisation des observations. La fiabilité des données est affaiblie et rend les résultats discutables. A.C. Berry (1976) a étudié la morphologie dentaire de diverses populations européennes, notamment du sud-ouest et nord-ouest de l’Angleterre, des îles Shetland et de l’Allemagne. Après avoir calculé des mesures de divergence entre les différents ensembles, les distances obtenues ont montré une bonne similarité avec la proximité géographique des différents groupes. P. Kirveskari (1978) s’est intéressé à la morphologie dentaire de la communauté lapone Skolt indiquant une origine européenne plutôt qu’asiatique, ce qui été aussi suggéré par les analyses sérologiques. G.R. Scott et A.A. Dahlberg (1982) ont étudié la morphologie dentaire de différentes tribus d’Indiens du sud-ouest américain qui se regroupent en quatre familles linguistiques. Leurs résultats montrent un fort parallélisme entre les données dentaires et linguistiques. Une autre étude a confirmé la similitude entre les données de la morphologie dentaire et la linguistique, il s’agit du travail de G.R. Scott et al. (1983), lesquels ont analysé un important échantillon d’Indiens Pima dont les relations familiales étaient connues. L’étude de A.J. Perzigian (1984) a comparé les résultats de données dentaires morphologiques et métriques. Il a étudié 42 échantillons répartis sur une fourchette chronologique très large (de la préhistoire à aujourd’hui). Selon lui, l’emploi de la morphologie pour distinguer les distances biologiques entre les populations est préférable à l’utilisation de la métrique. En effet, ses résultats montrent que les différences constatées pour la métrique dentaire peuvent être expliquées par des changements techno-culturels ou alimentaires, en revanche les données morphologiques dentaires permettent de regrouper les populations selon leur répartition géographique. C.R. Nichol (1990) a travaillé à la fois sur les données métriques et non métriques dentaires dans une perspective de compréhension des mécanismes de transmission de ce type de variables. Son travail a concerné l’étude de la dentition d’Indiens Pima de l’Arizona. Les résultats concernant la recherche d’affinités entre les différents groupes observés montrent une grande proximité des données dentaires (métriques et morphologiques) avec celles provenant de la linguistique. T. Higa et al. (2003) ont observé les variations morphologiques dentaires de différents ensembles asiatiques, plus particulièrement en s’intéressant aux populations de l’île Ryukyu. Ils ont effectué des analyses intra- et interpopulationnelles en obtenant des résultats comparables aux données fournies par la génétique. Différentes études ont exposé l’utilité des traits non métriques dentaires en tant qu’indicateurs des distances biologiques entre populations. Les comparaisons entre les différents indicateurs ont permis d’asseoir la valeur de la morphologie dentaire dans les études interpopulationnelles quelle que soit son échelle de différenciation. Les correspondances entre les distances dentaires et génétiques sont généralement plus que satisfaisantes. Les concordances entre la morphologie dentaire et la proximité géographique ainsi que la linguistique sont aussi très élevées, les différentes analyses ayant démontré de grandes analogies entre ces différents types de données. Lorsque les résultats ne sont pas en accord, cela est toujours expliqué par des problèmes méthodologiques qui se résument, la plupart du temps, par des problèmes d’observation et de réplicabilité des données. De plus, les études ayant porté sur des comparaisons entre les données métriques et non métriques ont montré que souvent ces dernières ont un meilleur pouvoir discriminant et semblent davantage refléter le déterminisme génétique tant recherché dans les études entre populations. 95 Chapitre 4 4.5. L’essentiel en bref Au terme de ce chapitre nous avons vu que les traits dentaires sont un outil intéressant pour appréhender les relations entre populations. D’une part, la dentition est l’un des éléments les plus résistants du squelette, donc souvent le mieux conservé. De l’autre, les traits non métriques dentaires peuvent être observés sur le vivant, ainsi la recherche sur leur déterminisme génétique est facilité puisqu’on peut directement étudier des sujets apparentés ou des jumeaux. Le référentiel à l’actualiste trouve ici toute son importance. Les progrès de la science ont aussi contribué à améliorer la compréhension des mécanismes et des processus impliqués dans l’odontogénie. • D’abord, les recherches ont pu établir que le développement dentaire, et en particulier celui de la morphologie dentaire, est soumis à un fort contrôle génétique. Plusieurs travaux ont montré que le développement de la dentition est contrôlé par un certain nombre de gènes qui agissent à divers endroits et interviennent à des moments distincts de sa formation. Si aucun trait ne suit un mode de transmission simple, ils ne sont pas aussi complexes que ceux des modèles de transmission polygénique pour lesquels interviennent une multitude de gènes. De plus, si on ne peut nier une influence probable de facteurs environnementaux, on ne peut pas non plus contester le fait que leurs effets sont moindres. Finalement, les études basées sur le déterminisme génétique ont montré que la plupart de ces caractères semblent détenir une grande composante héréditaire. • Ensuite, nous avons vu que la fiabilité du système d’observation est un élément essentiel dans la validation des données. En effet, l’absence de standardisation des données peut parfois conduire à des résultats totalement différents. La subjectivité de l’observation a souvent été un élément déterminant dans la critique de l’étude des traits dentaires. Mais à l’heure actuelle, il est possible de contourner cet obstacle, d’une part, en utilisant les plaques de référence et des définitions précises des différentes variables, et de l’autre, en éliminant les traits qui poseraient d’éventuels problèmes de standardisation en testant non seulement sa propre concordance, mais aussi celle entre différents observateurs. • Enfin, les études portant sur la valeur des traits non métriques dentaires en application dans des populations actuelles sont pleinement rassurantes. La comparaison des résultats de la morphologie dentaire avec différents estimateurs comme la proximité géographique, la linguistique ou encore des données génétiques, ont permis de conforter l’idée que ces variables sont de bons indicateurs des distances biologiques entre populations. Aujourd’hui, la recherche sur les traits non métriques dentaires se poursuit. Elle mène à une meilleure compréhension de ces variables. De plus, nous ne pouvons que nous réjouir des concordances des résultats entre l’anthropologie dentaire et la génétique puisqu’il semblerait que les dental data on northern Asia, southeast Asia and the Americas are generally in excellent agreement with those from single genes (Cavalli-Sforza et al. 1994, p. 317). Même si certains aspects demandent encore à être mieux maîtrisés, notamment les gènes responsables du développement de ces variables pour ne citer qu’un exemple, le déterminisme génétique qui sous-tend l’expression des traits dentaires permet de les utiliser lors d’études comparatives entre populations. 96 Chapitre 5 De la dent au trait S’il existe un certain nombre de variations morphologiques dentaires, toutes ne sont pas appropriées dans le cadre de notre travail. Ainsi, nous devons dans un premier temps sélectionner les traits dentaires qui fourniront des indications pertinentes sur l’histoire des populations que nous étudions. Ensuite, nous nous attacherons aux modalités d’acquisition des données. Finalement, nous évaluerons ici nos compétences tant dans la reconnaissance de la dentition que dans la détermination de l’expression des variations morphologiques retenues dans cette étude. 5.1. Les traits non métriques dentaires 5.1.1. La sélection des traits Cette étape est essentielle puisqu’elle a pour objectif la sélection des traits dentaires que nous observerons dans notre recherche. La littérature concernant les études des relations entre les populations à travers les variations morphologiques dentaires montre que les auteurs n’emploient pas régulièrement les mêmes traits. Certains traits sont souvent, et même toujours, étudiés, comme le tubercule de Carabelli, le protostyle ou encore les crêtes linguales marginales (trait plus connu sous l’appellation de dents en pelle) ; d’autres ne le sont pas forcément, voire pas du tout. Notre sélection s’appuie sur plus d’un siècle de recherches. Il est important de choisir des traits qui ne représentent pas différentes manifestations d’un caractère, ou au contraire, qui ne sont pas mêlés à divers traits, et ce, afin d’éviter toute redondance d’informations. Par exemple, certains chercheurs subdivisent les expressions du tubercule de Carabelli en deux variables distinctes. Le stade 1 (présence d’un orifice sur le sillon vestibulaire) est considéré comme un trait indépendant des autres expressions (présence de sillon - stade 2 - à une cuspide à part entière - stade 7 -). Le dessin du sillon, qui consiste à déterminer par des formes (en X, Y et +) la position des cuspides des molaires inférieures et leur zone de contact, est un trait souvent subdivisé par les chercheurs. Pour J.A. Sofaer et al. (1972a), l’expression en X et celle en Y sont des traits distincts et l’expression + représente l’absence de ces traits. Cette subdivision est fausse car la séquence du dessin du sillon est un continuum morphologique qui va de la forme en X, en passant par +, vers l’expression en Y. Le stade intermédiaire est alors considéré comme l’absence du trait, ce qui n’est pas correct d’un point de vue biologique (Nichol 1990). De ce fait, nous avons conçu une liste qui combine des caractères susceptibles d'être observés tant sur les dents permanentes (49 traits pour 334 observations possibles) que sur les déciduales (44 traits pour 196 observations possibles) (figures 29 et 30). La sélection de variables observées sur la dentition permanente se compose, d'une part, des traits retenus par le système ASU (Arizona State University) qui regroupe des caractères de format dichotomique ou gradué à l’observation standardisée. Ce système comprend également une série de moulages de référence des traits à expression graduée comme support supplémentaire aux définitions (Turner et al. 1991, Scott et Turner 1997). Ce système, largement adopté par la communauté scientifique travaillant sur les relations entre populations à travers l’étude de la dentition, propose des traits simples à regarder, possédant une certaine résistance à l’usure, une évolution lente et un dimorphisme sexuel généralement très bas (Turner et al. 1991). Cette liste est, d'autre part, complétée par une sélection de caractères appartenant au système FU-DTS (Freiburg University Dental Trait System). Ce système se compose de traits qui seraient hautement héréditaires (Alt et Türp 1995). Des études cliniques ont prouvé que les caractères présentés par le système FU-DTS sont rares dans les populations (Alt 1997). Ce système subdivise les traits en 97 Chapitre 5 anomalies et variantes qui seraient adaptées pour la détermination des relations entre les individus. Les traits retenus par ce système sont, pour la plupart, les mêmes que ceux présentés par le système ASU. La différence se situe au niveau des gradations, une large majorité de traits sont enregistrés sur la base de leur présence/absence. La liste mise en place pour les dents déciduales se compose, pour la plupart, de traits définis par K.D. Jørgensen (1956), K. Hanihara (1961, 1963) et P.W. Sciulli (1977). De plus, une partie des traits enregistrés sur les molaires permanentes, appartenant aux systèmes ASU et FU-DTS, sont observés sur les secondes molaires déciduales. Ces dernières, très proches morphologiquement de leurs homologues permanentes, devraient en toute cohérence exprimer les mêmes traits. Nous décidons, dans la mesure du possible, de garder les mêmes définitions et gradations proposées par les auteurs ayant établi les expressions des différents traits. Un long travail d’apprentissage s’impose pour les assimiler et les appliquer adroitement. Caractère observé Absence congénitale (congenital absence) Nombre de radicules (radical number) Racine appendiciforme (radiculae appendiciformes) Nombre de racines de la canine Nombre de racines des prémolaires (premolar root number) Racine de Tomes (Tomes root) Nombre de racines des molaires (molar root number) Racine paramolaire (radix paramolaris) Racine idiopathique (idiopathische radices) Racine de Carabelli (radix Carabelli) Racine de Citroën (radix Citroen) Racine entomolaire (radix entomolaris) Degré de rotation (winging) Convexité vestibulaire (labial convexity) Sillon traversant (interruption groove) Tubercule distal (akzessorische höckerchen) Cingulum vestibulaire (cingulum) Crêtes vestibulaires marginales (double-shoveling) Crêtes linguales marginales (shoveling) Tubercule dentaire (tuberculum dentale) Gouttière vestibulaire (talon cusp) Crête accessoire distale (canine distal accessory ridge) Crête mésiale (canine mesial ridge) Odontome (odontome) Cuspides accessoires (accessory cusp) Variation des cuspides linguales (lingual cusp variations) Extension de l'émail (enamel extensions) Parastyle (parastyle) Trait de Carabelli (Carabelli's trait) Métacone (metacone) Hypocone (hypocone) Métaconule (metaconule) Tubercule accessoire distal (distal accessory tubercle) Tubercule mésial paracone Tubercule mésial accessoire (mesial accessory tubercle) Protoconule (protoconule) Tubercule lingual paracone (lingual paracone tubercle) Pli cuspide mésiolingual (deflecting wrinkle) Fosse précuspidale (anterior fovea) Crête trigonide intermédiaire (mid trigonid crest) Crête trigonide distale (distal trigonid crest) Nombre de cuspides (cusp number) Dessin du sillon (groove pattern) Protostylide (protostylid) Hypoconulide (hypoconulid) Entoconulide (entoconulid) Métaconulide (metaconulid) Tubercule paracone (tuberculum paracone) Tubercule de Citroën (tuberculum Citroen) Application I2, P2, M3, I1, P2, M3 Toutes les dents I, C, P CP1, P2 P1 Molaires Molaires Molaires M1, M2, M3 M1 , M2, M3 M1, M2, M3 I1 I1, I 2 I1, I 2 I1, I 2 I1, I 2 I, C, P I, C, P I1, I 2, CI1, I 2, CCanines CPrémolaires P1, P2 P1, P2 P1, P2, M1 , M2, M3 M1, M2, M3 M1, M2, M3 M1, M2, M3 M1, M2, M3 M1, M2, M3 M1, M2, M3 M1, M2, M3 M1, M2, M3 M1, M2, M3 M1, M2, M3 M1 M1 M1, M2, M3 M1, M2, M3 M1, M2, M3 M1, M2, M3 M1, M2, M3 M1, M2, M3 M1, M2, M3 M1, M2, M3 M1, M2, M3 M1, M2, M3 Système d’enregistrement Système ASU (modifié) Système ASU Système FU-DTS Système ASU Système ASU Système ASU Système ASU (modifié) Système FU-DTS Système FU-DTS Système FU-DTS Système FU-DTS Système FU-DTS Système ASU (modifié) Système ASU Système ASU Système FU-DTS Système FU-DTS Système ASU Système ASU Système ASU (modifié) Système FU-DTS Système ASU Système ASU Système ASU Système ASU Système ASU Système ASU Système ASU Système ASU Système ASU Système ASU Système FU-DTS Système ASU Système FU-DTS Système FU-DTS Système FU-DTS Système FU-DTS Système ASU Système ASU Système ASU (modifié) Système ASU Système ASU Système ASU Système ASU Système ASU Système ASU Système ASU (modifié) Système FU-DTS Système FU-DTS (modifié) Figure 29 : Liste des traits non métriques dentaires observés sur la dentition permanente. La première colonne correspond au nom du trait suivi, entre parenthèses, du terme le plus souvent employé dans la littérature, la deuxième colonne désigne les dents sur lesquelles le caractère est observé et la dernière présente les principaux systèmes d’enregistrement. 98 De la dent au trait Lors de la sélection des variables, un trait dentaire a posé de sérieux problèmes quant à sa définition, car elle diffère selon les auteurs. Il s’agit du métaconule ou cuspide 5 que nous avons choisi parmi le système ASU. Ce trait est traditionnellement décrit comme un tubercule accessoire situé occlusalement sur le sillon marginal distal (Harris et Bailit 1980, Townsend et al. 1986, Turner et al. 1991), entre le métacone et l’hypocone. Pour K. Alt (1997), le métaconule et la cuspide 5 ne sont pas des termes synonymes pour définir une même variante mais deux traits distincts. Selon lui, le métaconule est un tubercule sur le sillon oblique, et la cuspide 5 est un tubercule sur le sillon marginal distal qui est aussi appelé tubercule accessoire distal. Il nous faut choisir entre ces différentes définitions et trouver s’il y a ou non une confusion des définitions parmi les auteurs. Nous estimons qu’une majorité de chercheurs appliquant une définition n’est pas une raison suffisante pour la considérer comme exacte. A travers une quête minutieuse d'informations qui permettraient d’éclaircir ces différences, nous trouvons finalement l’article répondant à nos interrogations. E. Kanazawa et al. (1990) étudient les différentes cuspides accessoires présentes sur les molaires supérieures. Ces auteurs confirment la confusion des définitions autour de ces deux termes. Ainsi, les définitions dont il faut tenir compte correspondent à celles proposées par K. Alt (1997). Seuls ces deux traits ont posé au départ un réel souci de compréhension de ce qu’il fallait voir. Caractère observé Application Nombre de radicules (radical number) Toutes les dents Extension de l'émail de la surface vestibulaire (enamel line) Toutes les dents Dilacération (dilaceration) i1, i2 Racine appendiciforme (radiculae appendiciformes) i, c Nombre de racines incisive et canine (incisors, canine root number) i, c Racine de la canine (canine root section) cNombre de racines molaires (molar root number) molaires Racine paramolaire (radix paramolaris) molaires Racine idiopathique (idiopatische radices) molaires Forme des incisives (incisors incisal border) incisives Racine de Carabelli (radix Carabelli) m1, m 2 Racine de Citroën (radix Citroen) m1, m 2 Racine entomolaire (radix entomolaris) m1, m 2 Gouttière vestibulaire (talon cusp) i1, i2 Crêtes linguales marginales (shoveling) i1, i2 , cTubercule dentaire (tuberculum dentale) i1, i2 , cDouble tubercule lingual (double lingual tubercle) cDéveloppement des cuspides (cusp development) m1 Crête du métacone m1 Parastyle (parastyle) m1, m 2 Trait de Carabelli (Carabelli's trait) m2 Métacone (metacone) m2 Hypocone (hypocone) m2 Métaconule (metaconule) m2 Tubercule accessoire distal (distal accessory tubercle) m2 Tubercule mésial paracone (mesial paracone tubercle) m2 Tubercule mésial accessoire (mesial accessory tubercle) m2 Protoconule (protoconule) m2 Tubercule lingual paracone (lingual paracone tubercle) m2 Développement des cuspides (cusp number) m1, m 2 Protostylide (protostylid) m1, m 2 Fosse précuspidale (anterior fovea) m2 Distostylide fissuré (split distostylid) m2 Crête trigonide intermédiaire (mid trigonid crest) m2 Crête trigonide distale (distal trigonid crest) m2 Pli mésiolingual (deflecting wrinkle) m2 Hypoconulide (hypoconulid) m2 Entoconulide (entoconulid) m2 Métaconulide (metaconulid) m2 Tubercule paracone (tuberculum paracone) m2 Tubercule de Citroën (tuberculum Citroen) m2 Fosse postérieure m2 Sillon trigonide (trigonid furrow) m2 Proéminence cervicale (cervical prominence) m2 Système d’enregistrement Système ASU Jørgensen (1956) (modifié) Jørgensen (1956) Système FU-DTS Jørgensen (1956) Jørgensen (1956) (modifié) Système ASU (modifié) Système FU-DTS Système FU-DTS Jørgensen (1956) (modifié) Système FU-DTS Système FU-DTS Système FU-DTS Système FU-DTS Hanihara (1963) Système ASU Jørgensen (1956) Hanihara (1963) (modifié) Jørgensen (1956) Système ASU Hanihara (1963) Système ASU Système ASU Système FU-DTS Système ASU Système FU-DTS Système FU-DTS Système FU-DTS Système FU-DTS Hanihara (1963) (modifié) Système ASU Système ASU Jørgensen (1956) Système ASU (modifié) Hanihara (1961) Système ASU Système ASU Système ASU Système ASU (modifié) Système FU-DTS Système FU-DTS (modifié) Jørgensen (1956) (modifié) Jørgensen (1956) Jørgensen (1956) Figure 30 : Liste des traits non métriques dentaires observés sur la dentition déciduale. La première colonne correspond au nom du trait suivi, entre parenthèses, du terme le plus souvent employé dans la littérature, la deuxième colonne désigne les dents sur lesquelles le caractère est observé et la dernière présente les principaux systèmes d’enregistrement. 99 Chapitre 5 Nous avons apporté quelques changements dans les gradations de certains traits proposées par les différents systèmes d’enregistrement. Nous avons effectué trois types de modification : la fusion de deux variables, une simplification des stades et une réorganisation de l’échelle d’expression. Seules deux variables ont fait l’objet d’une fusion. • Le premier trait fusionné est l’absence congénitale. Nous avons fusionné l’absence congénitale et la réduction en taille de la dent (peg-shaped tooth) des incisives et des molaires car il existe probablement un continuum de la réduction en taille d’une dent jusqu’à son absence. La variable est observée sur la dentition permanente uniquement. • La deuxième variable est le nombre de racines des molaires. Nous avons intégré le pyramidalisme - racine en forme de cône unique - dans la première expression du nombre de racines des molaires car nous avons l’impression qu’en les enregistrant séparément, nous obtiendrions deux fois la même information pour le premier stade (une racine). La variable est enregistrée sur les dentitions permanente et déciduale. Lorsque les gradations ne sont pas assez claires, nous avons préféré les simplifier. Ceci doit nous permettre d’éviter des erreurs d’appréciation dont nous avions conscience dès le départ. Ainsi, deux variables observées sur les deux dentitions ont été dichotomisées. • Le système ASU distingue pour l’enregistrement de la crête trigonide intermédiaire crête séparant les cuspides 1 et 2 - une gradation en 2 sous-stades (stades 1A et 1B). Nous optons pour un enregistrement simplifié basé uniquement sur la présence/absence du trait. • Le système FU-DTS adopte pour l’observation du tubercule de Citroën - tubercule se situant sur la surface linguale de la cuspide mésiolinguale - une gradation en 4 stades : des microformes (stade 1) au développement d’un grand tubercule (stade 4). Nous optons pour un enregistrement simplifié basé uniquement sur la présence/absence du trait. Nous avons également redéfini, pour quatre variables, observées sur la dentition permanente pour la première et sur la dentition déciduale pour les trois autres, des expressions qui ne nous semblent pas assez claires pour que nous réussissions à les observer. Nous choisissons délibérément de fusionner les stades qui présentent des différences trop fines et subtiles, en ne gardant que des groupes distincts et facilement observables. 100 • Le système ASU adopte pour le degré de rotation des incisives centrales supérieures une gradation en 4 stades selon la position de ces dernières. Deux sous-stades sont néanmoins présents pour la rotation bilatérale - incisives rotationnées mésiolingualement - (stades 1A : angle formé supérieur à 20° et 1B : angle inférieur à 20°). Nous optons pour un enregistrement simplifié en fusionnant les deux stades de la rotation bilatérale en ne gardant que la rotation et en éliminant l’angle formé par les incisives. • Pour l’extension de l’émail de la surface vestibulaire - coulée d'émail qui se situe souvent entre les deux racines -, K.D. Jørgensen (1956) définit 6 stades selon l’ampleur et la forme de la coulée. Nous avons simplifié les gradations en regroupant les deux premiers en stade 1 et les deux suivants en stade 2 car les différences entre ces stades sont beaucoup trop fines pour être reproduites convenablement. • Pour la racine de la canine - détermination de la courbure de la racine -, K.D. Jørgensen (1956) propose une gradation en 4 stades. Nous avons simplifié les gradations regroupant les degrés 1 et 2 en stade 1, pour les mêmes raisons que pour le trait précédent. De la dent au trait • La forme des incisives, variante également définie par K.D. Jorgensen (1956), est décomposée en 5 stades. Nous rencontrons également un problème de définition, c’est pourquoi nous réorganisons les différents stades. Nous avons simplifié et avons regroupé les deux premiers en stade 1 et les deux suivants en stade 2. Certains auteurs observent plusieurs critères simultanément pour juger de l’expression de certains traits. Nous trouvons l’exercice relativement fastidieux, c’est pourquoi nous avons décidé pour un trait observé sur la dentition déciduale d’en simplifier l’observation. • Le nombre de cuspides des molaires déciduales comprend 6 stades chez P.W. Sciulli (1977). Il tient compte, d’une part, de la présence d’une cuspide et, d’autre part, de sa taille. Nous décidons, par souci d’objectivité, de ne compter que le nombre de cuspides, sans tenir compte en même temps de leur grandeur respective, dans la mesure où cela nous semble laborieux à réaliser. Nous réorganisons l’échelle d’expression de deux traits, observés sur la dentition permanente pour le premier et sur les deux dentitions pour le deuxième. Ces transformations ne modifient en rien les définitions proposées par les différents systèmes d’enregistrement, il s’agit simplement de numéroter en continu, sans adjonction de subdivisions, les expressions de ces traits. • Le système ASU propose pour le tubercule dentaire - tubercule se situant dans la région cingulaire de la surface linguale - une gradation en 6 expressions, l’avantdernière subdivisée en 2 stades « 5- » et « 5 ». Nous choisissons une numérotation en continu, ainsi notre stade 5 correspond au stade 5- du système ASU, le 6 au 5 et le 7 au 6. • Le système ASU distingue pour le métaconulide - septième cuspide des molaires inférieures - une gradation en 4 stades : des microformes correspondant au stade 1 qui est subdivisé en 2 stades - 1A et 1B- au développement d’une cuspide à part entière (stade 4). Nous optons pour un enregistrement simplifié sans subdivision du premier stade avec les mêmes définitions mais une gradation en continu avec les mêmes définitions que celles d’ASU (stades 1 à 5). Tous les autres traits sélectionnés sont enregistrés selon les définitions mises en place par les différents systèmes et auteurs. 5.1.2. Les différents types de données dentaires Comme nous l’avons vu précédemment, la liste que nous avons définie permet de réaliser 334 observations par l’enregistrement de 49 variables pour la dentition permanente et 196 observations pour 44 variables pour la dentition déciduale. Les dents de l’arcade supérieure sont légèrement plus sollicitées par rapport à celles de l’arcade inférieure, respectivement 32 et 27 traits observés. Pour la dentition déciduale, 26 variables sont enregistrées par arcade29. Les dents antérieures, par leur morphologie peu compliquée et variée, ne proposent qu’un nombre restreint de variations morphologiques. Cela ne concerne pas les incisives et les canines supérieures définitives car, au même titre que les molaires, elles montrent des variations plus marquées, du moins au niveau de leur couronne. Les variations morphologiques dentaires sélectionnées se manifestent et s’enregistrent de diverses manières. Les figures 31 et 32 synthétisent les différents types de données que nous allons observer. 29 Les totaux ne correspondent pas au nombre de traits observés par dentition. Certaines variables comme le nombre de radicules, le nombre de racines des prémolaires et des molaires ou encore la crête accessoire distale... sont enregistrées sur les isomères mais pas nécessairement avec les mêmes gradations. Désormais, nous listerons et numéroterons séparément les mêmes variantes observées sur les deux arcades. Ainsi, les totaux s’élèvent à 59 variables observées sur la dentition permanente et 52 traits observés sur la dentition déciduale. 101 Chapitre 5 Nom du trait 1 Absence congénitale P2 1 Absence congénitale M3+I2 2 Nombre de radicules 3 Racine appendiciforme 4 Nombre de racines prémolaires 5 Nombre de racines molaires 6 Racine de Carabelli 7 Racine paramolaire 8 Racine idiopathique 9 Degré de rotation 10 Convexité vestibulaire 11 Sillon traversant 12 Tubercule distal 13 Cingulum vestibulaire 14 Gouttière vestibulaire 15 Crêtes linguales marginales I1+C 15 Crêtes linguales marginales I2 16 Tubercule dentaire I1 16 Tubercule dentaire I2+C 17 Crêtes vestibulaires marginales 18 Crête mésiale 19 Crête accessoire distale 20 Cuspides accessoires 21 Odontome 22 Extension de l'émail 23 Parastyle 24 Trait de Carabelli 25 Métacone 26 Hypocone 27 Métaconule 28 Tubercule distal accessoire 29 Tubercule mésial paracone 30 Tubercule mésial accessoire 31 Protoconule 32 Tubercule lingual paracone 33 Absence congénitale P2 33 Absence congénitale M3+I1 34 Nombre de radicules 35 Racine appendiciforme 36 Nombre racines canines 37 Racine de Tomes 38 Nombre racines molaires 39 Racine de Citroën 40 Racine paramolaire 41 Racine entomolaire 42 Racine idiopathique 43 Crêtes linguales marginales 44 Crêtes vestibulaires marginales 45 Crête accessoire distale 47 Variation des cuspides linguales 46 Odontome 48 Pli cuspide mésiolinguale 49 Fosse précuspidale 50 Crête trigonide intermédiaire 51 Crête trigonide distale 52 Nombre de cuspides 53 Dessin du sillon 54 Protostylide 55 Hypoconulide 56 Entoconulide 57 Métaconulide 58 Tubercule paracone 59 Tubercule de Citroën Code utilisé numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique alphabétique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique alphabétique numérique numérique numérique numérique numérique numérique Type dichotomique gradué ordonné gradué ordonné dichotomique gradué ordonné gradué ordonné dichotomique dichotomique dichotomique gradué et non ordonné gradué ordonné non gradué et non ordonné dichotomique dichotomique dichotomique gradué ordonné gradué ordonné gradué ordonné gradué ordonné gradué ordonné gradué ordonné gradué ordonné dichotomique dichotomique gradué ordonné gradué ordonné gradué ordonné gradué ordonné gradué ordonné dichotomique gradué ordonné dichotomique dichotomique dichotomique dichotomique dichotomique gradué ordonné gradué ordonné dichotomique gradué ordonné gradué ordonné gradué ordonné dichotomique dichotomique dichotomique dichotomique gradué ordonné gradué ordonné gradué ordonné gradué ordonné dichotomique gradué ordonné gradué ordonné dichotomique dichotomique gradué ordonné non gradué et non ordonné gradué ordonné gradué ordonné gradué ordonné gradué ordonné dichotomique dichotomique Expression présence/absence développement/absence développement absence/présence nombre nombre absence/présence absence/présence absence/présence position développement position absence/présence absence/présence absence/présence absence/développement absence/développement absence/développement absence/développement absence/développement absence/développement absence/développement absence/présence absence/présence absence/développement absence/développement absence/développement absence/développement absence/développement absence/présence absence/développement absence/présence absence/présence absence/présence absence/présence présence/absence développement/absence développement absence/présence nombre absence/développement nombre absence/présence absence/présence absence/présence absence/présence absence/développement absence/développement absence/développement développement et position absence/présence développement absence/développement absence/présence absence/présence nombre position absence/développement absence/développement absence/développement absence/développement absence/présence absence/présence Gradation 0-3 0,1,2,3 1,2,3,4,5,6,7,8 0-1 1,2,3 1,2,3,4 0-1 0-1 0-1 1,2,3,4 0,1,2,3,4 0, M, D, M, Méd 0-1 0-1 0-1 0,1,2,3,4,5,6 0,1,2,3,4,5,6,7 0,1,2,3,4 0,1,2,3,4,5,6,7 0,1,2,3,4,5,6 0,1,2,3 0,1,2,3,4,5 0-1 0-1 0,1,2,3 0,1,2,3,4,5,6 0,1,2,3,4,5,6,7 0,1,2,3,4,5,6 0,1,2,3,4,5,6 0-1 0,1,2,3,4,5 0-1 0-1 0-1 0-1 0-1 0,1,2,3 1,2,3,4,5,6,7,8 0-1 1,2 0,1,2,3,4,5 1,2,3 0-1 0-1 0-1 0-1 0,1,2,3 0,1,2,3,4,5,6 0,1,2,3,4,5 0,1,2,3,4,5,6,7,8,9 0-1 0,1,2,3 0,1,2,3,4 0-1 0-1 4,5,6 Y,+, X 0,1,2,3,4,5,6,7 0,1,2,3,4,5 0,1,2,3,4,5 0,1,2,3,4,5 0-1 0-1 Figure 31 : Le type des traits non métriques dentaires observés sur la dentition permanente. La première colonne correspond au nom de la variable précédé par un chiffre qui se réfère au numéro du trait sur la fiche d’enregistrement, la deuxième colonne désigne le format du code d’enregistrement, la troisième l’échelle de mesure du système, la quatrième présente la signification des expressions codées et la dernière les gradations que nous avons retenues. 102 De la dent au trait Nom du trait 1 Nombre de radicules 2 Extension de l'émail 3 Dilacération 4 Racine appendiciforme 5 Nombre de racines i et c 6 Racine de la canine 7 Nombre de racines molaires 8 Racine de Carabelli 9 Racine paramolaire 10 Racine idiopathique 11 Forme des incisives 12 Gouttière vestibulaire 13 Crêtes linguales marginales 14 Tubercule dentaire 15 Déveleppoment cuspides 16 Crête du métacone 17 Parastyle 18 Trait de Carabelli 19 Métacone 20 Hypocone 21 Métaconule 22 Tubercule distal accessoire 23 Tubercule mésial paracone 24 Tubercule mésial accessoire 25 Protoconule 26 Tubercule lingual paracone 27 Nombre de radicules 28 Extension de l'émail 29 Racine appendiciforme 30 Nombre de racines i et c 31 Nombre de racines molaires 32 Racine de Citroen 33 Racine paramolaire 34 Racine idiopathique 35 Racine entomolaire 36 Forme des incisives 37 Double tubercule lingual 38 Développement cuspides m1 38 Développement cuspides m2 39 Protostylide 40 Fosse précuspidale m1 40 Fosse précuspidale m2 41 Distostylide fissuré 42 Crête trigonide intermédiaire 43 Crête trigonide distale 44 Pli mésiolingual 45 Hypoconulide 46 Entoconulide 47 Métaconulide 48 Tubercule paracone 49 Tubercule de Citroen 50 Fosse postérieure 51 Sillon trigonide 52 Proéminence cervicale Code utilisé numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique numérique Type gradué ordonné gradué ordonné gradué et non ordonné dichotomique gradué ordonné gradué et non ordonné gradué ordonné dichotomique dichotomique dichotomique gradué non ordonné dichotomique gradué ordonné gradué ordonné gradué ordonné gradué ordonné gradué ordonné gradué ordonné gradué ordonné gradué ordonné dichotomique gradué ordonné dichotomique dichotomique dichotomique dichotomique gradué ordonné gradué ordonné dichotomique gradué ordonné gradué ordonné dichotomique dichotomique dichotomique dichotomique gradué non ordonné dichotomique gradué ordonné gradué ordonné gradué ordonné dichotomique gradué ordonné dichotomique dichotomique dichotomique gradué ordonné gradué ordonné gradué ordonné gradué ordonné dichotomique dichotomique dichotomique dichotomique dichotomique Expression développement absence/développement position absence/présence nombre forme nombre absence/présence absence/présence absence/présence forme absence/présence absence/développement absence/développement nombre absence/développement absence/développement absence/développement absence/développement absence/développement absence/présence absence/développement absence/présence absence/présence absence/présence absence/présence développement absence/développement absence/présence nombre nombre absence/présence absence/présence absence/présence absence/présence forme absence/présence nombre nombre développement absence/présence absence/développement absence/présence absence/présence absence/présence absence/développement absence/développement absence/développement absence/développement absence/présence absence/présence absence/présence absence/présence absence/présence Gradation 1,2,3,4,5,6,7,8 0,1,2,3 0,1,2 0-1 1,2,3 0,1,2 1,2,3 0-1 0-1 0-1 1,2,3 0-1 0,1,2,3 0,1,2,3,4,5,6 2,3,4 0,1,2 0,1,2,3,4,5,6 0,1,2,3,4,5,6,7 0,1,2,3,4,5,6 0,1,2,3,4,5,6 0-1 0,1,2,3,4,5 0-1 0-1 0-1 0-1 1,2,3,4,5,6,7,8 0,1,2,3 0-1 1,2,3 1,2,3 0-1 0-1 0-1 0-1 1,2,3 0-1 3,4,5 4,5,6 0,1,2,3,4,5,6,7 0-1 0,1,2,3,4 0-1 0-1 0-1 0,1,2,3 0,1,2,3,4,5 0,1,2,3,4,5 0,1,2,3,4,5 0-1 0-1 0-1 0-1 0-1 Figure 32 : Le type des traits non métriques dentaires observés sur la dentition déciduale. La première colonne correspond au nom de la variable précédé par un chiffre qui se réfère au numéro du trait sur la fiche d’enregistrement, la deuxième colonne désigne le format du code d’enregistrement, la troisième l’échelle de mesure du système, la quatrième présente la signification des expressions codées et la dernière les gradations que nous avons retenues. Il existe deux manières de coder les observations. Le code peut être numérique ou alphabétique. La première option est celle rencontrée pour la majorité des traits dentaires. Le code alphabétique est employé seulement pour deux variables (le sillon traversant et le dessin du sillon). Nous verrons par la suite que le type de code utilisé a finalement peu d’importance contrairement à la signification de 103 Chapitre 5 l’expression qui, elle, est fondamentale. Néanmoins, les traitements statistiques, préliminaires ou analytiques, imposent une gradation numérique, impliquant une transformation des codes alphabétiques. Ainsi, les variables morphologiques s’expriment de façon très différentes. Une manifestation peut être limitée à son absence ou à sa présence, comme par exemple la présence d’une racine accessoire ou un tubercule. Elle peut également montrer différents stades de développement, comme la plupart des traits concernant les cuspides des molaires. Cependant un caractère peut toujours être présent, on s’attachera alors à observer son développement, son nombre, sa forme ou encore sa position. Nous avons donc regroupé en catégories les différents types de données dentaires : • Le trait dichotomique : Il s’agit de l’alternative la plus élémentaire car la variable est observée selon son absence ou sa présence. • Le trait gradué ordonné : La variable est observée sur la base de son développement. Le caractère peut être absent ou non, néanmoins il montre dans sa présence différents stades d’expression. • Le trait gradué et non ordonné : La variable est enregistrée sous forme numérique mais ses différentes expressions ne sont pas d’ordre développemental. Ce sont des variantes qui traduisent une forme (comme l’aspect de la section de la racine de la canine) ou une position (comme le degré de rotation des incisives). L’ordre des gradations n’est pas un reflet de l’évolution de la manifestation. • Le trait non gradué et non ordonné : La variable est enregistrée sous forme alphabétique mais est de même type que celle graduée et non ordonnée. Seuls deux traits sont de ce type, il s’agit du sillon traversant et du dessin du sillon. Une description complète des traits se trouve en annexe 3 pour les dents permanentes et en annexe 4 pour les dents déciduales. Ces deux catalogues comprennent une fiche descriptive de chaque trait. Le titre correspond au numéro du trait sur la fiche d’enregistrement manuel, à son nom en français et à sa traduction anglaise (parfois latine ou allemande selon l’auteur ayant défini le trait). Chaque fiche s’articule ensuite en huit rubriques. La première concerne les différents termes rencontrés dans la littérature. Ensuite, nous présentons non seulement les références utiles pour la description des traits, un champ présentant les dents sur lesquelles le trait est observé, une brève description du trait, mais aussi le système d’enregistrement sélectionné et les gradations retenues. Puis, un champ est destiné aux commentaires éventuels. La fiche se termine généralement par une représentation du trait sous forme de photographies ou de croquis. Une ultime rubrique indique s’il existe une plaque de référence pour les différentes expressions des traits. Cette phase, concernant la sélection et la mise en place des définitions et des différentes gradations, doit nous permettre des observations systématiques faciles à reproduire. 104 De la dent au trait 5.2. L’acquisition des données Nous allons ainsi observer les traits non métriques dentaires sélectionnés sur l’ensemble des populations des régions retenues. Il faut encore mettre en place un système d’acquisition des données. Par expérience, nous savons que les manipulations d’observation du matériel et leur enregistrement sous forme informatisée en simultané ne sont pas des plus aisées. De ce fait, nous devons concevoir une fiche manuelle d’enregistrement des données. Ensuite, il nous faut créer une fiche informatisée sous forme de base de données où nous retranscrirons toutes les observations en vue des futures analyses. 5.2.1. La création d’une fiche d’enregistrement manuel Nous avons créé deux fiches d’enregistrement reproduisant les dentitions permanente et déciduale facilitant l’acquisition des données (annexe 5). Ces fiches se divisent en trois parties distinctes. • La première concerne les informations générales de l’individu observé. Nous notons les informations suivantes : sous la rubrique site le nom du site, sous la rubrique n°site son numéro (en référence à la base de données des sites étudiés en annexe 1), sous la rubrique fiche un numéro que nous attribuons à l’individu (de 1 à n par région), sous numéro nous indiquons le numéro de l’individu ou de la tombe (selon les informations spécifiques à chaque site). De plus, des indications sur les dents elles-mêmes sont spécifiées, comme le type de dents auquel nous avons affaire, leur localisation (maxillaire ou mandibule) et la présence de dents n’appartenant pas à l’individu. • La deuxième partie concerne les traits dentaires proprement dits où nous enregistrons les expressions des variantes observées. La première colonne donne, outre le nom du trait observé, le numéro de la variable en référence au catalogue en annexes 1 et 2 et les gradations possibles. Les différents quadrants de la dentition sont disposés en vue occlusale de manière à faciliter l’enregistrement des données. • La dernière partie est une zone réservée à d’éventuelles observations en cours d’enregistrement. Les fiches manuelles sont remaniées lors de l’enregistrement des dents isolées. Nous séparons alors les fiches par isomères, ceci faisant partie d’un processus naturel et écologique d’économie de papier. Nous obtenons donc une fiche dent à la place de la fiche individu (annexe 6). 5.2.2. La création d’une base de données Nous avons conçu une base de données permettant la gestion de toutes les observations à l'aide d'un fichier unique (annexe 7). Ce formulaire de saisie, que nous avons créé avec le logiciel Access, permet non seulement le traitement et la présentation simplifiée de l’information, mais consent également le transfert des données dans d’autres systèmes d’analyse. A l’intérieur de ce fichier, nous répartissons les informations entre plusieurs contenants appelés tables. Celles-ci reproduisent fidèlement les fiches d’enregistrement manuel. Le masque d’enregistrement est divisé en 5 parties. Nous devons procéder à cette subdivision puisque le logiciel Excel n’admet pas plus de 256 variables par fichier et nous avons 530 variables dentaires par individu. Le passage par ce logiciel est une phase transitoire, mais obligatoire, pour accéder aux programmes de gestion statistique. La première partie se rapporte aux informations sur l’individu et fera le lien avec les 4 autres parties qui correspondent aux maxillaires et mandibules des dentitions permanentes et déciduales. L’avantage d’un tel formulaire de saisie, contrairement à celui manuel, est de permettre d’avoir toutes les informations sous les yeux en même temps. Bien entendu, aucun processus naturel et écologique d’économie de giga ou de méga n’est mis en place pour la base de données ! 105 Chapitre 5 5.3. L’évaluation de la concordance intra- et interobservateurs Cette partie du travail est une étape nécessaire pour assurer une certaine légitimité à nos données. Une longue phase d’apprentissage est nécessaire afin de pouvoir atteindre un niveau de compétence acceptable. Il faut non seulement posséder une connaissance irréprochable de la dentition (identification de la dent, latéralisation…), mais il est également impératif de maîtriser les définitions des différents traits dentaires et de réussir à les observer. En pratiquant des observations morphologiques, un chercheur doit sans cesse prendre des décisions. Ainsi, nous allons évaluer non seulement la concordance dans la détermination des dents entre deux chercheurs, mais également la concordance des modes d’observations des traits non métriques dentaires entre nos propres observations et celles d’un second observateur. 5.3.1. L’évaluation de la concordance dans la détermination de la dentition Il n’est pas ordinaire de rencontrer dans la littérature des recherches prenant en considération l’évaluation de la détermination dentaire. Il s’agit, somme toute, d’un élément important dans la mesure où un chercheur n’est pas toujours amené à travailler sur des collections présentant uniquement des dents sur arcade. Et dans le cadre de notre recherche, nous avons travaillé à la fois sur des échantillons composés d’individus que sur des séries constituées essentiellement de dents isolées. L’idée d’estimer le taux d’erreur dans la détermination des dents de l’une des séries de notre corpus a été réfléchie après l’enregistrement des données du dolmen des Peirières (Villedubert, Aude, France). Il s’agit d’un ensemble constitué presque exclusivement de dents isolées. Ce monument possède au moins deux occupations, l’une datant du Néolithique final, l’autre du Campaniforme. Nous avons été en mesure d’effectuer ce test grâce à la collaboration de Coralie Demangeot, doctorante en anthropologie à l’Université de Bordeaux. Elle a également étudié la série de Villedubert dans le cadre de son travail de doctorat portant sur les méthodes de calcul du NMI (nombre minimum d’individus) (Demangeot en cours). Nous avons ainsi pu comparer nos données, même si, dans le cadre de notre travail, la démarche consiste principalement à vérifier la numérotation/cote de chaque dent en vue de leur restituer une bonne attribution culturelle (table 1). Nous avons simplement effectué des statistiques descriptives sans autre prétention que d’obtenir un bilan des discordances dans la détermination des dents entre deux observateurs. traits dentaires - TD - J. Desideri nombre minimum d’individus - NMI - C. Demangeot 2763 3875 Figure 33 : Nombre total de dents enregistrées par observateur. Nous constatons dans un premier temps que le nombre total de dents n’est pas le même pour chaque observateur (figure 33). Une différence de plus de 1100 dents est observée. Il s’agit avant tout de dents indéterminées (marquage et/ou type de dent non défini) dont le nombre s’élève à 899 qui sont prises en compte pour l’étude du NMI et à 35 (marquage non défini) pour les traits dentaires - TD. Le nombre peu élevé de cas indéterminés pour les TD s’explique par le fait que seules les dents sur lesquelles des traits non métriques ont pu être observés ont été retenues. En éliminant des totaux les dents inclassables, le nombre total de dents observées pour le NMI s’élève à 2976 et à 2728 pour les TD, par conséquent la différence ne devrait être plus que de 248 cas. Mais, étonnamment, des différences auxquelles nous n’avions pas pensé sont bel et bien présentes. La figure 34 synthétise les divers cas de figures rencontrés. détermination commune concordante détermination commune discordante détermination NMI uniquement détermination TD uniquement détermination TD autre site 106 2368 219 389 107 34 Figure 34 : Composition des différentes observations de la série de Villedubert. De la dent au trait Les déterminations communes concordantes se retrouvent dans 2368 cas et le nombre d’attributions communes discordantes s’élève à 219. L’étude du NMI présente 387 dents supplémentaires qui n’ont pas été enregistrées pour les traits non métriques dentaires. Nous n’avons malheureusement pas le détail de ces déterminations puisque nous travaillons sur des tableaux qui nous ont été transmis par Coralie Demangeot et n’avons plus le matériel sous les yeux. Une bonne partie de ces dents doivent faire partie de sujets inutilisables dans notre travail comme les bourgeons dentaires, les racines isolées ou encore les dents fortement usées. Il est également probable que certaines dents qui auraient pu être enregistrées aient échappé à notre attention. Le problème inverse est aussi présent. Nous avons examiné 106 dents que nous n’avons pas retrouvées dans la base de données du NMI. Nous n’avons pas les moyens de vérifier s’il est possible que des dents aient également échappé à l’attention de Coralie Demangeot ou alors si elles peuvent appartenir à un autre ensemble (la numérotation homogène ne nous permet pas de trancher). Elles seront donc éliminées afin d’éviter d’introduire d’éventuelles erreurs dans l’interprétation. Nous avons encore enregistré 34 dents appartenant à l’aven de la Boucle (Corconne, Gard, France), sépulture collective datant du Néolithique final, qui étaient stockées par mégarde dans les mêmes cartons que ceux du dolmen des Peirières. Celles-ci ont été immédiatement éliminées de notre base de données. Nous nous sommes par la suite intéressée aux 219 déterminations communes discordantes en précisant leur origine et leur répartition (figure 35). erreur de notation avec erreur latéralisation avec erreur inter-arcade avec erreur intradistrict avec erreur interdentition avec erreur interdistrict erreur de latéralisation avec erreur inter-arcade avec erreur intradistrict avec erreur interdentition avec erreur interdistrict erreur notation 79 13 6 67 3 5 13 erreur interarcade avec erreur latéralisation avec erreur intradistrict avec erreur interdentition avec erreur interdistrict erreur notation erreur intradistrict avec erreur latéralisation avec erreur inter-arcade avec erreur interdentition avec erreur interdistrict erreur notation 13 3 4 76 5 4 6 erreur interdistrict 11 avec erreur latéralisation avec erreur inter-arcade avec erreur intradistrict avec erreur interdentition erreur notation erreur interdentition 3 avec erreur latéralisation avec erreur inter-arcade avec erreur intradistrict avec erreur interdistrict erreur notation Figure 35 : Décompte des déterminations communes discordantes selon leur origine et leur répartition. Nous avons donc réparti les discordances en plusieurs catégories distinctes et constatons qu’il est également possible de rencontrer deux types différents de discordance sur une même dent. • les erreurs de notation Il s’agit des erreurs de transcription du marquage lors de l’observation des données. Ce type d’erreur s’élève à 79 cas. Elles sont la conséquence d’une mauvaise lecture ou transcription du numéro attribué lors de la fouille. Ces dernières sont de petites dimensions et de forme peu adéquate au marquage. L’exercice de déchiffrage se révèle souvent ardu et, sans surprise, induit des erreurs qui peuvent avoir des effets importants. Nous devons dans ce travail attribuer une phase culturelle aux dents d’après leur numérotation. Il est important de ne pas introduire d’erreur de ce type dans notre échantillon, il est donc nécessaire d’éliminer celles présentant ce genre de discordance. On les trouve parfois combinées à des erreurs de latéralisation (13 cas) et à des erreurs intradistrict (6 cas). 107 Chapitre 5 • les erreurs de latéralisation Il s’agit d’un désaccord d’attribution du côté de la dent. Le désaccord du choix de l’antimère s’élève à 67 cas. Ce problème se retrouve en majorité sur les incisives latérales supérieures (7 cas) et les secondes prémolaires inférieures (11 cas). Même si le nombre de discordances est relativement bas, nous sommes étonnée de cette différence puisqu’il ne s’agit pas de dents posant généralement des problèmes de latéralisation (du moins en ce qui nous concerne) et si nous avions à prédire quelle dent pourrait être plus facilement sujette à des erreurs de latéralisation, nous aurions spontanément sélectionné les incisives inférieures. • les erreurs interarcade Il s’agit du désaccord de l’attribution de l’arcade dentaire de la dent. La confusion entre les isomères s’élève à 13 cas. Les canines ont été le plus souvent sujettes à ce type d’erreur (8 cas). La morphologie très proche entre les isomères de ce district peut mener à ce genre de confusion. Somme toute, ce genre d’erreur n’est pas abondamment représenté. • les erreurs intradistrict Il s’agit de la confusion de la position de la dent dans le district dentaire. Ce type d’erreur est le plus fréquent, on le retrouve à 76 reprises. Les molaires, qui ont le plus souvent fait l’objet de ce type de divergence, sont des dents difficilement distinguables entre elles (au sein du même district) (32 cas). Les prémolaires supérieures (19 cas), que l’on reconnaît principalement par la hauteur respective des cuspides vestibulaire et linguale ainsi que les incisives inférieures (12 cas), le deviennent également dès la présence d’usure. • les erreurs interdistrict Il s’agit d’un désaccord dans l’attribution de la classe dentaire d’une dent, désaccord s’élevant à 11 cas. La morphologie de la dentition suit un continuum des incisives aux molaires, c’est pourquoi il est possible parfois de confondre le district d’une dent avec l’une des classes adjacentes. Les erreurs ne dépassent jamais un désaccord de plus d’un champ dentaire, donc les confusions sont de type incisive-canine (5 cas) ou canineprémolaire (5 cas) ou encore prémolaire-molaire (1 cas). • les erreurs interdentition Il s’agit de la confusion de l’attribution du type de dentition. Ce type de divergence est relativement rare. Trois mêmes cas ont été observés. Il s’agit de la confusion entre la seconde molaire déciduale et la première molaire permanente. Elles sont en tous points morphologiquement similaires. Généralement trois éléments permettent de distinguer la molaire temporaire de la définitive : la forme plus évasée de ses racines, sa couronne plus bombée et sa petite taille. Notre principal intérêt ici correspond en l’attribution culturelle correcte des sous-ensembles de ce monument, celui secondaire à l’évaluation de la concordance dans la détermination de la dentition entre deux chercheurs. Nous n’avons pas réalisé de tests statistiques, cependant les discordances, que l’on ne retrouve pas en abondance, ne semblent pas subir de biais systématiques et pourraient refléter un taux de variation naturelle auquel nous pourrions nous attendre. De plus, la morphologie des différents types de dent n’est pas figée, ainsi il est possible de se tromper en toute bonne foi. Malgré tout, en enregistrant des traits bien précis sur les différentes dents, nos erreurs ne devraient pas être les plus importantes. C’est pourquoi, nous décidons d’éliminer toutes les dents qui présentent des discordances de notation (79 cas) et de garder nos déterminations pour les divergences du type de dent. Pour aller au bout de cette démarche, nous devrions idéalement reprendre en collaboration avec Coralie Demangeot point par point les divergences et en faire leur analyse. Nous ne pouvons pas l’achever ici, mais il serait intéressant de le réaliser dans le futur pour mieux appréhender les variations du nombre de déterminations, davantage que les divergences de déterminations elles-mêmes. 108 De la dent au trait 5.3.2. L’évaluation de la concordance des modes d’observation des traits non métriques dentaires Nous avons vu lors du chapitre précédent qu’il est impératif d’acquérir une standardisation des modes d’observation afin de garantir la reproduction des résultats. De nombreux chercheurs soulignent le caractère subjectif de l’évaluation de l’expression des caractères morphologiques dentaires, que ce soit entre observateurs ou par un même chercheur à différents moments de la recherche. Une concordance parfaite de l’attribution des expressions des différentes variables d’un échantillon observé à plusieurs reprises relève de l’utopie. Néanmoins, il est possible de réduire les différences par l’utilisation de plaques de référence, de définitions précises et de l’expérience pour qu’elles n’aient aucune incidence significative sur les fréquences populationnelles. Pour C.R. Nichol et C.G. Turner (1986), les difficultés varient selon les observateurs, c’est pourquoi il est important d’évaluer la concordance des modes d’observation dans une recherche. Nous testons non seulement les variations de nos propres modes d’observation, mais également en confrontation avec un second chercheur30. Un échantillon a été sélectionné au sein du matériel étudié comportant plus d’une trentaine d’individus dont les dents permettent l’enregistrement d’un nombre important d’observations. Nous souhaitons, par la même occasion, reprendre des individus observés au début de notre travail afin d’avoir une idée de l’évolution personnelle des différentes phases d’enregistrement31. L’échantillon se compose d’individus provenant des nécropoles de Barmaz I et II (Collombey-Muraz, Valais, Suisse). Seuls 42 traits dentaires observés sur la dentition permanente font l’objet d’une évaluation puisque les variables retenues par notre second observateur ne correspondent pas toutes à celles que nous enregistrons dans ce travail. Trois séries d’observations sont ainsi disponibles pour cette analyse (table 2) : • O1a : fichier des observations réalisées lors de la première session d’enregistrement. • O1b : fichier des observations effectuées lors de la seconde série d’enregistrement. • O2 : fichier qui comprend la session d’enregistrement de comparaison (observations de S. Eades). Il s’agit ici de comparer non seulement nos deux séries d’enregistrement (O1a/O1b), mais également chacune de nos sessions avec celle de comparaison (O1a/O2 et O1b/O2). Nous allons présenter les résultats des concordances entre observateurs, même s’il ne s’agit pas d’un élément essentiel pour la poursuite de cette recherche. L’intérêt est de quantifier les divergences d’appréciation entre deux chercheurs ayant effectué un apprentissage commun des traits non métriques dentaires. Cela semble, à première vue, une situation idéale où les différences devraient être moindres. Dans un premier temps, nous évaluerons les divergences absolues présentes entre les différentes sessions d’enregistrement. Dans un deuxième temps, nous avons choisi, parmi les huit valeurs utilisées par C.R. Nichol et C.G. Turner (1986) dans leur étude concernant l'examen des erreurs intra- et interobservateurs, cinq tests qu’ils ont mis en place pour saisir la variabilité des éventuelles discordances. La première valeur consiste à déterminer les divergences d’enregistrement d’un trait pendant une session qui n’est pas enregistré pendant une session différente. Ensuite, nous nous intéressons aux discordances de gradations. Pour ce faire, nous abordons différents aspects pour comprendre les causes et les conséquences de ces divergences : le taux total de discordance, la variation de plus d’un stade de différence et l’erreur moyenne brute. Enfin, nous effectuerons un test de similarité des observations entre les sessions d’enregistrement. Il sera intéressant de comparer cas par cas nos propres résultats aux leurs et à ceux de S. Eades qui a également effectué ce type de tests (Eades 2003). 30 Suzanne Eades, qui a réalisé une thèse au Département d’anthropologie et d’écologie de l’Université de Genève sur l’héritabilité des traits dentaires (Eades 2003), a eu l’amabilité d’observer les 35 individus sélectionnés afin que nous puissions également tester les erreurs entre observateurs dans ce travail. 31 Une année s’est écoulée entre les deux phases d’enregistrement. 109 Chapitre 5 Cette étape a pour objectif de déterminer que les divergences d’appréciation ne sont pas trop importantes et qu’elles n’ont pas d’impact sur les échantillons étudiés. Selon les résultats, il faudra peut-être redéfinir les gradations qui posent problèmes et si nécessaire envisager l’élimination de certaines variables. 5.3.2.1. L’évaluation de la discordance absolue Cette première valeur consiste à estimer les discordances brutes entre deux séries d’observations. Il s’agit simplement de quantifier toute déviation ou écart de jugement indépendamment de leur origine. Les pourcentages individuels par trait et par session concernant cette mesure se trouvent en table 3. session étendue moyenne O1a-O1b 0-42.05% 13.48% O1a-O2 O1b-O2 0.97-76.92% 3.88-60.00% 36.08% 30.00% Figure 36 : Evaluation des divergences absolues. La figure 36 présente l’étendue et la moyenne générale des variations d’enregistrement d’une session à l’autre calculées à partir des trois fichiers d’observation. Les valeurs se situent entre 0% et 42.05% pour les discordances intra-observateurs et entre 0.97% et 76.92% pour celles interobservateurs, avec des moyennes respectives à 13.48% et entre 30% et 36.08%. Ces résultats, concernant toutes les déviations possibles d’enregistrement, expriment bien les difficultés que l’on peut avoir à reproduire fidèlement les observations faites sur un même échantillon. D’une manière générale, la série intra-observateur montre que les variables enregistrées sur les dents supérieures présentent les divergences les plus élevées. Sur l’ensemble des traits, 9 variables font l’objet d’une réplication parfaite. Il s’agit avant tout de traits dichotomiques et de variables dont l’expression traduit une position bien précise comme le sillon traversant (T11), où l’on note la position d’un sillon sur la surface linguale des incisives. 14 traits se situent en dessous de 10% de discordance absolue. Les autres traits ont des étendues très diverses. Il ne s’agit pas uniquement de traits gradués, on y trouve également des traits dichotomiques et des variables à expression non développementale. Il faudra alors en mesurer l’importance et déterminer leur origine car, selon les causes, les incidences sur les analyses peuvent être plus ou moins importantes. La situation des deux séries interobservateurs est très explicite puisque sur l’ensemble des deux sessions aucun trait ne présente une reproduction identique des observations. En effet, seuls 2 traits se placent en dessous du seuil de 10% de discordance absolue. Il s’agit des crêtes linguales marginales (T44, valeur entre 0.97% et 4.85%) et des crêtes vestibulaires marginales (T45, valeur à 3.88%) observées sur les incisives inférieures. Nous ne sommes pas réellement étonnée puisque ces deux traits facilement enregistrables présentent très peu de variabilité. Pour le reste, les déviations les plus importantes ne sont pas en relation avec le nombre d’expression des variables. Il sera fort intéressant d’affiner la perception de ces résultats par les prochains tests qui permettront sans aucun doute de mieux comprendre l’organisation des difficultés. 5.3.2.2. L’évaluation de la concordance de l’observation des traits Le nombre de variables observables par trait dans un échantillon peut différer selon la session et l’observateur. On ne tient pas compte du stade attribué mais uniquement de l’observabilité ou non d’un trait. C.R. Nichol et C.G. Turner (1986) nomment cette valeur observed on one session only percentage. Les variations rencontrées peuvent êtres dues à divers facteurs. Il peut s’agir au départ d’une réticence à enregistrer une variable, réticence qui peut s’estomper par la pratique. La situation inverse est également possible. L’usure dentaire est aussi un facteur paralysant l’enregistrement d’une variable et l’expérience peut modifier, en limitant ou en étendant, l’observabilité de celle-ci. 110 De la dent au trait L’observation d’un trait peut donc se révéler difficile et c’est alors qu’il faut choisir un seuil d’acceptation au-delà duquel une variable est considérée comme difficilement réplicable. C.R. Nichol et C.G. Turner (1986) ont défini un seuil à 10%, valeur également suivie par S. Eades (2003). Les pourcentages individuels par trait et par session concernant cette mesure se trouvent en table 3. session étendue moyenne O1a-O1b 0-41.91% 10.62% Nichol1-Nichol2 (Nichol et Turner 1986) 0-33.30% 7.10% Nichol1-Nichol3 (Nichol et Turner 1986) 0-29.60% 6.70% Nichol2-Nichol3 (Nichol et Turner 1986) 0-39.60% 7.70% Eades1-Eades2 (Eades 2003) 0-78.60% 12.20% O1a-O2 O1b-O2 0-61.54% 0-50.53% 26.60% 21.03% Nichol1-Turner (Nichol et Turner 1986) 2-64.30% 14.70% Nichol2-Turner (Nichol et Turner 1986) 2-50.00% 12.10% Nichol3-Turner (Nichol et Turner 1986) 0-51.90% 13.10% Figure 37 : observed on one session only percentage Pourcentage de l’enregistrement ou non d’un trait entre deux sessions. La figure 37 présente l’étendue et la moyenne générale de l’enregistrement ou non d’un trait d’une session à l’autre calculées à partir des trois fichiers d’observation. Nos valeurs, qui se situent entre 0% et 41.91% pour les discordances intra-observateurs et entre 0% et 61.54% pour celles interobservateurs, se placent dans les marges proposées par les différents auteurs, avec des moyennes légèrement supérieures à celles indiquées dans les autres études. Pour les discordances intra-observateurs et sur l’ensemble des traits observés, 12 d’entre eux montrent une observabilité identique. L’origine des écarts pour les autres variables montre qu’avec l’expérience nous les avons enregistrées plus facilement. 18 traits se situent au-delà du seuil d’acceptation. Ces traits sont en majorité enregistrés sur les dents supérieures. Il s’agit, avant tout, de traits rares comme le tubercule distal (T12, valeur à 40.91%) ou encore les doubles gouttières (T14, valeur à 39.47%). Nous avons eu, au départ, des difficultés à observer ce type de trait tant leur présence est exceptionnelle au sein de nos populations. Les autres traits concernés sont ceux qui se situent sur les faces d’implantation de l’usure comme la surface occlusale des molaires (le trait de Carabelli - T24, le métacone - T25 et l’hypocone - T26) ou encore les faces linguales des incisives (les crêtes linguales marginales - T15). Le problème n’est pas d’échouer dans l'appréciation de la présence d’un trait, mais dans l’attribution de son expression. Ainsi, en cas d’incertitude d’évaluation correcte d’un stade à une manifestation sur une dent présentant de l’usure, nous finissons souvent par ne pas l’enregistrer. En général, ces deux problèmes se règlent par la pratique et l’expérience. Sans grand étonnement, une nette majorité de variables dépasse plus ou moins largement le seuil d’acceptation pour les deux séries concernant les discordances interobservateurs. Nos sessions (O1a et 1b) ont une observabilité plus élevée par rapport à celle de comparaison (O2). La plupart des traits concernés sont ceux qui n’ont montré aucune variation d’enregistrement intra-observateur, indiquant probablement une plus grande prudence de la part du second observateur. 5.3.2.3. L’évaluation de la discordance des graduations enregistrées Cette valeur vise à estimer les discordances entre les stades proposés pour une même manifestation. Il s’agit donc de tester la capacité d’attribution et de reproduction d’un degré d’expression. C.R. Nichol et C.G. Turner (1986) nomment la valeur inverse variant scoring percentage. Ils ont défini un seuil d’acceptation de divergence absolue à 10%, valeur également adoptée par S. Eades (2003). Les pourcentages individuels par trait et par session concernant cette mesure se trouvent en table 3. 111 Chapitre 5 session étendue moyenne O1a-O1b 0-23.81% 3.34% Nichol1-Nichol2 (Nichol et Turner 1986) 0-65.90% 26.20% Nichol1-Nichol3 (Nichol et Turner 1986) 0-76.70% 29.40% Nichol2-Nichol3 (Nichol et Turner 1986) 0-70.00% 25.90% Eades1-Eades2 (Eades 2003) 0-96.40% 22.60% O1a-O2 O1b-O2 0-57.78% 0-46.14% 13.25% 11.02% Nichol1-Turner (Nichol et Turner 1986) 0-80.50% 31.70% Nichol2-Turner (Nichol et Turner 1986) 2-79.10% 29.00% Nichol3-Turner (Nichol et Turner 1986) 0-66.10% 0-51.9% 30.10% Figure 38 : variant scoring percentage Evaluation des discordances de gradations d’un trait d’une session à l’autre. La figure 38 présente l’étendue et la moyenne générale des variations d’évaluation de la gradation d’un trait d’une session à l’autre calculées à partir des trois fichiers d’observation. Nos valeurs, tant pour les discordances intra-observateurs que pour celles interobservateurs, sont considérablement inférieures à celles qui sont indiquées dans les autres études, d’environ 20% pour la moyenne et pour la marge supérieure de l’étendue. Pour les discordances intra-observateurs, les traits observés sur les dents inférieures ont une divergence d’appréciation relativement basse, inférieure à 12% (sauf T47) avec une moyenne à 2%. Les différences pour les observations réalisées sur les dents supérieures sont légèrement plus élevées avec une moyenne à 5% avec des valeurs comprises entre 0% et 23.81%. Les écarts totaux ont une moyenne de 3.34%. Plus précisément, 19 traits montrent une concordance parfaite entre les deux séries d’enregistrement. Les variantes qui entrent dans cette catégorie sont soit des traits rares, soit des variables dichotomiques. Pour le reste, 9 traits se placent au-dessous du seuil d’acceptation, dont 4 ont une discordance très basse (environ 1%). Les variables restantes, au nombre de 4, se rangent en dessus du seuil. Il s’agit du cingulum vestibulaire (T13, valeur à 11.11), du métacone (T25, valeur à 13.85), de l’hypocone (T26, valeur à 23.81) et de la variation des cuspide linguales (T47, valeur à 11.76). Ces variables font partie des traits présentant plus de quatre expressions différentes. Pour les deux séries concernant les discordances interobservateurs, nous remarquons au premier abord que les difficultés qui se dessinent sont les mêmes que celles intra-observateurs, mais bien plus accentuées. Ceci est un élément très rassurant, cette situation traduit une certaine logique dans la difficulté de réplication des observations. Les discordances se situent aux mêmes endroits avec une présence de taux plus élevés. Ceci implique une constance dans la difficulté d’attribution de gradations de certaines variantes seulement. Les difficultés se situent, à nouveau, sur les traits possédant plus de trois expressions dans leur variabilité. En général, les traits à expression dichotomique présentent aussi de bons résultats. Ainsi, 16 variables pour la première comparaison entre observateurs et 15 pour la seconde dépassent le seuil d’acceptation. 5.3.2.4. Le pourcentage d’attribution de plus d’un stade de différence entre deux sessions Cette valeur est relativement importante et consiste à calculer le taux de discordance de plus d’un stade dans l’attribution d’une gradation. En effet, C.R. Nichol et C.G. Turner, qui nomment cette valeur >1 grade variant scoring, estiment que because grading standards of the nature used for the examination of morphological characters divide a continuous range of expression into discrete classes [...] that disagreements of one grade represents a natural difficulty in the ability to consistently classify individuals [...] whereas disagreements of two or more grades are due either to imprecise observational technique, recording error, or difficulties with standard itself (Nichol et Turner 1986, p. 300). Ces auteurs ont défini un seuil d’acceptation de divergence absolue à 10%, valeur également adoptée par S. Eades (2003). Ce test ne concerne pas les variables exprimées dichotomiquement, c’est pourquoi cette valeur est calculée 112 De la dent au trait sur les 27 traits gradués seulement. Les pourcentages individuels par trait et par session concernant cette mesure se trouvent en table 3. session étendue moyenne O1a-O1b 0-13.49% 1.79% Nichol1-Nichol2 (Nichol et Turner 1986) 0-25.70% 26.20% Nichol1-Nichol3 (Nichol et Turner 1986) 0-38.30% 29.40% Nichol2-Nichol3 (Nichol et Turner 1986) 0-25.00% 25.90% Eades1-Eades2 (Eades 2003) 0-50.00% 6.50% O1a-O2 O1b-O2 0-31.25% 0-36.84% 7.72% 6.04% Nichol1-Turner (Nichol et Turner 1986) 0-80.50% 31.70% Nichol2-Turner (Nichol et Turner 1986) 0-79.10% 29.00% Nichol3-Turner (Nichol et Turner 1986) 0-66.10% 0-51.9% 30.10% Figure 39 : >1 grade variant scoring Evaluation de l’attribution de plus d’un stade de différence. La figure 39 présente l’étendue et la moyenne générale des variations d’attribution de plus d’un stade de différence d’une session à l’autre calculées à partir des trois fichiers d’observation. Nos valeurs se situent dans les marges proposées par les différents auteurs, avec des moyennes et des marges supérieures nettement inférieures à celles indiquées dans les autres études. Ce type d’erreur n’est pas abondamment présent dans la série intra-observateur. En effet, sur l’ensemble des variables, 13 n’en présentent pas et 13 autres se situent en dessous du seuil d’acceptation. Seule une variable, l’hypocone (T26), se situe au-dessus avec une valeur à 13,49%, valeur qui n’est pas des plus alarmantes. La moyenne est très basse (moins de 2%) signifiant que les écarts d’appréciation de plus d’un stade sont très rares. Ces résultats ont une importance fondamentale, puisqu’ils révèlent qu’aucun trait ne fait l’objet de divergences de standardisation graves. Les résultats des comparaisons entre observateurs sont également très bons. Seuls 6 traits pour la première série inter-obervateur et 5 variables pour la seconde se placent au-dessus du seuil des 10%. Les moyennes et les marges supérieures sont également très basses. 5.3.2.5. L’erreur moyenne brute Cette valeur, que C.R. Nichol et C.G. Turner (1986) désignent comme net mean grade difference ou NMGD, définit l’erreur moyenne d’une gradation exprimée en pourcentage. Il s’agit d’une valeur brute qui prend en compte la direction des divergences entre deux sessions d’enregistrement. Une valeur positive indique que des expressions plus importantes ont été attribuées lors de la seconde série d’observation. Une mesure négative indique, à l’opposé, que des gradations plus élevées ont été constatées dans la première série. Une erreur moyenne proche de zéro révèle que les écarts sont aléatoires et non directionnels. Ces auteurs ont défini un seuil d’acceptation de l’erreur moyenne brute qui se traduit par une pondération en rapport avec le nombre d’expressions possibles par trait32. Pour S. Eades ces expressions maximales sont rarement atteintes pour des populations européennes (Eades 2003, p. 43) et l’emploi du seuil d’acceptation pondéré proposé par les auteurs américains ne se justifie pas et elle a ainsi choisi un seuil d’acceptation différent s’élevant à ± 20%. Ce seuil semble relativement haut, c’est pourquoi nous décidons de garder un seuil à 10%. Celui-ci, plus restrictif, est induit par les différentes valeurs calculées précédemment. Nos divergences sont bien moins importantes que celles présentées par C.R. Nichol et C.G. Turner (1986) et par S. Eades (2003) et l’utilisation de leurs seuils n’est pas légitime. 32 Le seuil d’acceptation pondéré proposé par C.R. Nichol et G.R. Turner (1986) consiste à multiplier le nombre de stades maximum par variables par un seuil de 0.05. 113 Chapitre 5 Nous avons décidé de tester ici uniquement l’erreur moyenne brute de nos propres observations, l’intérêt de ce genre d’informations est moindre dans une perspective interobservateur. Les pourcentages individuels concernant cette mesure se trouvent en table 4. session étendue moyenne O1a-O1b -6.41 - 13.13% 0.67% Nichol1-Nichol2 (Nichol et Turner 1986) -21.70 - 68.60% 10.50% Nichol1-Nichol3 (Nichol et Turner 1986) -58.50 - 91.4% 1.20% Nichol2-Nichol3 (Nichol et Turner 1986) -57.60 - 44.60% -9.70% Eades1-Eades2 (Eades 2003) -81.00 - 100% -0.30% Figure 40 : net mean grade difference Evaluation de l’erreur moyenne brute. La figure 40 présente l’étendue et la moyenne générale des erreurs moyennes brutes. Nos résultats se placent dans les marges proposées par les différents auteurs avec une moyenne et une étendue bien inférieures. 13 variables montrent une erreur moyenne positive, indiquant que nous leur avons attribué des gradations plus élevées lors de la seconde session d’enregistrement. Au contraire, 7 traits avec une erreur moyenne brute négative montrent que nous avons abaissé leur expression lors du second passage. La valeur de l’erreur moyenne des variables restantes est égale à zéro. 19 d’entre elles ne montraient pas de discordance de graduation. Leur calcul n’était pas nécessaire mais cela correspond finalement à une vérification des données entre les différents tests. La valeur nulle des 3 variables restantes, soit les crêtes linguales marginales (T15), les crêtes vestibulaires marginales (T17) des dents antérieures supérieures et le nombre de cuspides (T52), indiquent que les écarts ne sont pas directionnels. Seuls deux traits se placent légèrement en dessus du seuil d’acceptation. Il s’agit de la crête mésiale (T18, valeur à 10.91) et de la variation des cuspides linguales (T47, valeur à 13.13). Toutes les autres valeurs ne s’écartent que très légèrement de la moyenne indiquant que les différences constatées sont négligeables et aléatoires. 5.3.2.6. Le degré de similarité des distributions La dernière valeur mesurée est un test de similarité des distributions. Selon le type de données, nous devons effectuer des tests distincts. Ainsi, afin de vérifier le degré de similarité des variables dichotomiques, il est conseillé d’utiliser un test du Chi2 ; en revanche, la similitude entre les distributions des traits gradués est mesurée à l’aide d’un test de t (paired sample t test). Les mesures sont réalisées sur nos propres observations et non sur les données entre observateurs. Sur l’ensemble des traits, 15 sont de format dichotomique et 27 sont de format gradué. Le premier type de variables se réfère soit à des caractères rares, soit leur observation est simple et claire. De ce fait, dans la plupart des cas, il s’agit de variables à expression constante. Nous trouvons également des paires d’observations invariantes, il est donc inutile de tester ce type de variables. Nous nous attacherons donc aux variables graduées uniquement (test de t) et nous ne procéderons à aucun test du Chi2. Les tests de t, qui s’effectuent au seuil de 5%, comparent les moyennes obtenues par sessions d’observation (O1a et O1b) pour chaque trait. Les résultats concernant ces tests se trouvent en table 5. Nous déterminons les hypothèses de départ qui sont les suivantes : H0 = hypothèse nulle : les deux moyennes sont homogènes, aucun biais systématique d’observation ne peut être mis en évidence. H1 = hypothèse alternative : les moyennes présentent des différences significatives, les divergences entre les deux séries d’enregistrement dénotent probablement des difficultés de standardisation. 114 De la dent au trait Le test de t n’a pas pu être calculé sur l’ensemble des variables, car pour certaines d’entre elles l’erreur standard de la différence entre les moyennes est égale à 0. Ainsi, 15 caractères observés sur les dents supérieures et 8 traits sur les dents inférieures ont pu être testés. Seul un trait, la variation des cuspides linguales des prémolaires inférieures (T47, test de t=-2.19, dl=79, p=0.037) ne répond pas à l’exigence de l’hypothèse nulle (H0). Nous pourrions rejeter l’hypothèse nulle pour ce trait avec le risque de commettre une erreur de type I (conclure de manière erronée que des différences significatives de standardisation sont présentes pour ce trait). Néanmoins, en considérant le nombre limité de test (n=42) nous pouvons abaisser le seuil d’acceptation à 0.01 en référence à la correction de Bonferroni (seuil alpha divisé par nombre de comparaisons) généralement adoptée lorsque l’effectif des tests est petit. Avec ce nouveau seuil, la variation des cuspides linguales des prémolaires inférieures (T47) se situe au-dessus des paramètres d’acceptation. C’est pourquoi nous conservons l’hypothèse 0 pour toutes les variables avec le risque de commettre une erreur de type II (conclure de manière incorrecte qu’aucun biais systématique ne peut être mis en évidence pour ces différents traits). 5.3.2.7. La concordance de l’enregistrement des traits dentaires : un bilan Nous avons testé ici la concordance de l’enregistrement des traits dentaires entre nos propres observations et, dans une perspective plus descriptive, nos données ont été comparées à celles effectuées sur le même échantillon par un second observateur (S. Eades). Nous utilisons dans ce travail uniquement des données que nous avons enregistrées, c’est pourquoi les tests entre observateurs n’étaient pas indispensables pour notre recherche mais essentiellement informatifs. En ce qui concerne l’analyse de nos sessions d’enregistrement, les résultats des différentes valeurs calculées ont permis de mieux cerner les origines des divergences. L’évaluation de la discordance absolue a montré que 19 traits sur 42 présentaient des écarts d’appréciation de plus de 10%. Le choix du seuil pour cette valeur est arbitraire et descriptif car il faut préciser l’origine des divergences pour lui donner un sens. Ainsi, nous avons calculé deux valeurs permettant de définir deux types de discordance : celle de l’observabilité (évaluation de la discordance de l’observation des traits) et celle de la gradation (évaluation de la discordance des graduations) qui a été elle-même subdivisée afin d’affiner les causes des écarts (attribution de plus d’un stade de différence, l’erreur moyenne brute et le degré de similarité des distributions). Les discordances se situent avant tout au niveau de l’observabilité des données, où l’on trouve des valeurs comparables à celles des autres études avec des moyennes légèrement supérieures. Les résultats concernant les variations d’attribution de gradations sont pleinement rassurants. En effet, si plus de la moitié des traits testés présentent des divergences d’appréciation, seules 4 variables se placent audessus du seuil d’acceptation. En général, les écarts concernent tout au plus 1 stade de différence et sont d’ordre aléatoire et non directionnel. Les valeurs que nous obtenons pour ces différents tests sont systématiquement inférieures à celles présentées dans les autres études et de façon très nette. L’hypocone (T26) est un trait qui se place souvent en zone critique du seuil d’acceptation. Il semblerait que nous ayons attribué des valeurs plus élevées lors de la seconde session d’enregistrement, des valeurs qui sont comparables à celles que l’on constate en général sur leur homologue dans l’échantillon régional étudié. La variation des cuspides linguales des prémolaires inférieures (T47) est le seul trait à se situer légèrement au-dessus du seuil d’acceptation pour le calcul de l’erreur moyenne brute. Il s’agit en général des observations faites sur la première prémolaire, la seconde ne posant aucun problème. Sa valeur montre que nous avons également attribué des expressions plus élevées lors du deuxième passage. Le test du degré de similarité a mis en évidence des différences significatives pour cette variable à un seuil de 5%, mais en abaissant ce dernier à 1% (par correction de Bonferroni) il ne se distingue pas des autres variables. Nous sommes certaine de ne pas introduire au sein de nos analyses une variable pourvue d’un biais systématique puisqu’il s’agit d’une variable qui ne nous pose, en général, aucun problème d’observation. D’une manière générale, les évaluations entre chercheurs révèlent des difficultés similaires à celles rencontrées entre nos propres observations, mais plus prononcées. Ceci est un élément très rassurant, cette situation traduit une certaine logique dans la difficulté de réplication des observations. Les discordances se situent aux mêmes endroits avec une présence de taux plus élevés. Ceci implique une constance dans la 115 Chapitre 5 difficulté d’attribution de gradations de certaines variantes seulement. De plus, le second observateur enregistre une multitude de traits supplémentaires afin de tester leur héritabilité dans un corpus aux relations familiales connues. L’apport de nouveaux traits explique parfois le niveau de concordance très bas de certains pourcentages. L’appréciation des expressions des variables peut être influencée et modifiée par la présence de nouveaux traits à observer aux mêmes endroits. Les taux de discordance se placent toujours dans les marges d’erreurs présentées par C.R. Nichol et C.G. Turner (1986) et ordinairement avec des valeurs bien inférieures. Nous n’avons pas testé toutes les variables adoptées dans cette recherche, comme les traits enregistrés sur les racines ou sur les dents déciduales. Les variations morphologiques des racines ne sont pas des plus compliquées et se résument la plupart du temps à leur décompte ou à l’observation de leur présence. Les variables enregistrées sur la dentition temporaire sont pour la plupart également observées sur la dentition définitive. Néanmoins, nous décidons tout de même d'écarter 6 variables observées sur la dentition déciduale : • Le pli mésiolingual (T44 - dentition déciduale) Ce trait observé à la fois sur la première molaire définitive et la seconde molaire déciduale fait partie du système ASU. Il faut définir la variation de la morphologie de l'arrête médiale de la crête de la deuxième cuspide (mésiolinguale). Si l’observation de ce trait sur la dentition permanente ne pose pas réellement de problèmes, son enregistrement sur la dentition de lait est plus difficile. • La racine de la canine (T6 - dentition déciduale) Cette variable, qui a été mise en place par K.D. Jørgensen (1956), concerne la détermination de la courbure de la racine par l’observation de sa section à la hauteur cervicale. • La crête du métacone (T16 - dentition déciduale) Ce trait a été défini par K.D. Jørgensen (1956). Le métacone comporte habituellement trois crêtes. Il est possible que l’une d’entre elles soit absente et l’objectif est de déterminer laquelle. • Le distostylide fissuré (T41 - dentition déciduale) Ce caractère a été mis en place par K.D. Jørgensen (1956). Le distostylide des molaires inférieures peut être subdivisé par la présence d’un sillon (ou fissure). • La fosse postérieure (T50 - dentition déciduale) L’observation de ce trait, défini par K.D. Jørgensen (1956), consiste à déterminer la présence d’une fosse postérieure (distale) sur l’hypoconulide de la seconde molaire inférieure. Nous avions dès le départ simplifié l’observation de ce trait en le codant dichotomiquement, plutôt que d’évaluer les différentes expressions de son développement. • Le sillon trigonide (T51 - dentition déciduale) Cette dernière variable est définie également par K.D. Jørgensen (1956). Il s’agit de juger si le sillon mésiodistal principal de la seconde molaire inférieure se dédouble ou non à son extrémité mésiale. Au terme de cette partie concernant la standardisation des observations, nous estimons que nous nous retrouvons face à une situation reflétant la réalité imposée par la pratique de cette discipline. Si nous savions qu’il est impossible d’obtenir 100% de concordance dans des échantillons observés à plusieurs reprises, nous avons démontré que les différences rencontrées sont minimes et, après avoir éliminé les variables susceptibles d’introduire un bruit, voire un biais, au sein des analyses, nous pouvons poursuivre notre travail sereinement. 116 De la dent au trait 5.4. L’essentiel en bref Ce chapitre avait comme objectif la sélection des variables que nous utilisons dans ce travail, le mode d’acquisition et l’évaluation de la concordance des données. • Dans un premier temps, nous avons sélectionné 111 traits de format dichotomique ou gradué représentant 530 observations possibles. Pour la dentition permanente, nous avons retenu des traits proposés par le système ASU (Turner et al. 1991) et par le système FU-DTS (Alt 1997). Pour la dentition déciduale, nous avons choisi des traits définis par K. Hanihara (1963), P.W. Sciulli (1977) et K.D. Jorgensen (1956). De plus, une partie des traits enregistrés sur les molaires permanentes sera observée sur les secondes molaires déciduales. Nous avons retenu, dans la majorité des cas, les expressions proposées par les différents auteurs et systèmes d’enregistrement. 11 traits ont tout de même fait l’objet de modifications car les gradations présentées ont été jugées comme difficile à identifier. • Dans un deuxième temps, nous nous sommes intéressée au mode d’acquisition des données. Nous avons conçu, d’une part, des fiches d’enregistrement manuel et, de l’autre, un système de gestion informatisé permettant la manipulation aisée et le transfert rapide de données pour une utilisation dans différents logiciels. • Enfin, nous avons mesuré la concordance dans la détermination des dents et dans l’observation des traits non métriques dentaires. L’évaluation de la concordance dans la détermination de la dentition a été réalisée même si l’objectif principal de la démarche résidait en l’attribution culturelle correcte d’un échantillon constitué essentiellement de dents isolées (dolmen des Peirières, Villedubert, Aude, France). Nous avons pu mettre en évidence différents type d’erreurs (notation, latéralisation, interarcade, interdistrict ou encore interdentition). Les discordances ne se retrouvent pas en abondance. L’erreur la plus souvent rencontrée est celle du déchiffrage incorrect du numéro attribué lors de la fouille (79 cas), les dents présentant ce type d’erreur ont été éliminées. L’évaluation de la concordance des modes d’observations des traits non métriques dentaires a fourni des résultats intéressants. Les résultats intra-observateurs indiquent que les différences de gradations entre nos séances d’enregistrement sont minimes et par conséquent négligeables. Nous avons néanmoins éliminé 6 variables observées sur la dentition déciduale que nous avions jugées difficile à observer. En revanche, les différences entre observateurs sont plus importantes et permettent de confirmer la difficulté d’utiliser des données que l’on n’a pas collectées soi-même. 117 Chapitre 6 Le traitement préliminaire des données Cette phase de traitement préparatoire est essentielle, car il s’agit d’une étape permettant non seulement la vérification de la transcription informatique des données, mais également leur épuration et leur optimisation en vue des analyses statistiques. Dans un premier temps, il faut transformer les données alphabétiques en variables numériques afin d’être exploitées dans les analyses statistiques. Puis, nous décomptons les observations qui ont pu être effectuées par région. Ensuite, nous vérifions que les bases de données ne contiennent pas d’éventuelles erreurs de transcription. Nous nous attachons enfin aux expressions des différents traits mobilisables pour l’étude par l’élimination des traits non pertinents et la mise en évidence des variables à expression constante qui seront éliminées selon les analyses. Dans un second temps, toute une série de tests de corrélations est effectuée, servant à optimiser les données et à réduire au maximum les perturbations ou bruits affectant l’expression des traits dentaires. De ce fait, l’expression bilatérale des traits est vérifiée afin de pouvoir fusionner les antimères. Ensuite, nous examinons les relations entre les expressions des différents traits par le comportement d’une variable au sein d’une même classe dentaire ou encore d’un trait s’exprimant à travers plusieurs districts. Finalement, nous contrôlons si les échantillons contiennent des traits qui s’expriment en fonction du sexe des individus. 6.1. Les statistiques descriptives Cette première étape consiste ainsi à calculer des statistiques descriptives sur les données brutes afin d’obtenir différents types d’informations concernant non seulement le corpus étudié, mais aussi des données descriptives comme les moyennes, les écarts-types, les minimums et les maximums. 6.1.1. Le traitement des traits non numériques Afin de pouvoir traiter statistiquement les données, les expressions des variables enregistrées sous format alphabétique sont transformées en données numériques. Seuls deux traits font l’objet de cette conversion : • Le sillon traversant (T11) observé sur la face linguale des incisives supérieures permanentes (sillon parcourant le cingulum, franchissant souvent la jonction cément-émail en direction de la racine). Le sillon peut être positionné de 4 manières différentes. Le choix de la numérisation pour cette variable est totalement arbitraire. Ainsi, l’absence du trait est notée 0, la position mésiale (M) du sillon 1, la position distale (D) 2, la médiane (Méd) 3 et enfin la double position mésio-distale (MD) 4. • Le dessin du sillon (T53) observé sur les molaires inférieures permanentes (forme du sillon en rapport avec la position des cuspides). Pour C.R. Nichol (1990) la séquence du dessin du sillon est un continuum morphologique qui va de la forme en X (les cuspides mésiovestibulaire et distolinguale sont en contact), en passant par l’expression intermédiaire en + (les 4 cuspides sont en contact en un point), vers l’expression en Y (les cuspides mésiolinguale et distovestibulaire sont en contact). Nous avons donc opté pour une traduction de l’expression selon cet auteur avec Y = 0, + = 1 et X = 2. 119 Chapitre 6 6.1.2. Le décompte total des observations Nous profitons de cette étape pour estimer le nombre d’observations (le nombre d’entrées) réalisées pour chaque région étudiée. Les différentes régions totalisent près de 255’000 entrées réparties sur plus de 2’000 individus. Les détails pour chacune d’entre elles sont donnés par les figures 41 à 45. Dents permanentes maxillaire mandibule 61’809 33’087 28’722 Dents déciduales 4’279 maxillaire mandibule 1’900 2’379 Total 66’088 Figure 41 : Décompte des observations pour la Bohême. Dents permanentes maxillaire mandibule 13’002 7’564 5’438 Dents déciduales maxillaire mandibule 400 184 216 Total 13’402 Figure 42 : Décompte des observations pour la Hongrie. Dents permanentes maxillaire mandibule 87’886 49’917 37’969 Dents déciduales maxillaire mandibule 9’436 5’281 4’155 Total 97’322 Figure 43 : Décompte des observations pour le sud de la France. Dents permanentes maxillaire mandibule 19’201 10’243 8’967 Dents déciduales maxillaire mandibule 741 371 370 Total 19’951 Figure 44 : Décompte des observations pour le nord de l’Espagne. Dents permanentes maxillaire mandibule 48’894 25’074 23’820 Dents déciduales maxillaire mandibule 9’205 4’262 4’943 Total 58’099 Figure 45 : Décompte des observations pour la Suisse. 120 Pour la Bohême, le corpus se compose d’un peu plus de 500 individus. Les observations qui ont pu être réalisées sur cet ensemble s’élèvent à plus de 65'000 entrées (table 6), dont la très large majorité ont été effectuées sur la dentition permanente. Le nombre d’entrées pour la dentition déciduale n’est pas négligeable, nous verrons plus loin comment les intégrer dans les analyses. La Hongrie est la région qui présente le plus petit effectif, une centaine d’individus seulement. Les observations qui ont pu être réalisées sur cet ensemble s’élèvent à plus de 13'000 entrées (table 7). Elles reposent principalement sur la dentition permanente. En effet, seules 400 observations, qui ne sont pas suffisantes pour être utilisées dans les analyses, ont pu être effectuées sur la dentition déciduale. L’échantillon pour le sud de la France est le plus conséquent, avec près de 700 individus. Les observations qui ont pu être réalisées sur cet ensemble s’élèvent à près de 100'000 entrées (table 8), dont la très large majorité ont été effectuées sur la dentition permanente. Le nombre d’entrées pour la dentition déciduale n’est pas négligeable, nous verrons plus loin comment les intégrer dans les analyses. Pour le nord de l’Espagne près de 180 individus ont pu être observés. Le nombre d’enregistrements s’élève à près de 20'000 entrées (table 9). Ils reposent principalement sur la dentition permanente. En effet, seules 741 observations ont été réalisées sur la dentition déciduale. Elles ne sont donc pas suffisantes pour être mobilisées dans les analyses. Pour la Suisse environ 520 individus ont été enregistrés. Le nombre d’observations s’élève à plus de 58'000 entrées (table 10). La répartition par dentition est identique à celle des autres régions, mais, proportionnellement, les données appartenant à la dentition déciduale sont nettement supérieures ici. Elles feront aussi l’objet d’une discussion pour savoir comment les intégrer dans les analyses. Le traitement préliminaire des données 6.1.3. La vérification de la transcription Une vérification d’éventuelles erreurs de transcription lors du report des observations manuelles dans la base de données a été réalisée à l’aide des minimums et des maximums calculés dans la phase de statistiques descriptives (tables 6 à 10). Ainsi, différents types d’erreurs ont pu être mis en évidence : • Les erreurs de frappe Ces erreurs sont assez facilement détectables et peuvent être modifiées sans problème puisque les gradations deviennent incohérentes. Il s’agit principalement de doublon de la gradation (par exemple, 22 au lieu de 2). • La gradation aberrante Ce type d’erreur concerne en général des traits dont l’expression minimum est autre que zéro. Par exemple le nombre de racines où le stade minimum correspond à 1 que nous avons noté par mégarde 0. Ce genre de variables a fait l’objet d’une vérification rigoureuse. • Le décalage de la saisie informatisée Lors de la saisie informatisée des fiches d’enregistrement, il est arrivé qu’il y ait un décalage de l’information. Ces erreurs sont très facilement repérables puisque la majorité des entrées d’une fiche montrent des expressions aberrantes. Après plusieurs phases de vérification et de correction, nous avons tiré au hasard 200 fiches par région. Nous avons pu constater avec sérénité qu’elles présentaient une absence totale d’erreurs. 6.1.4. La pertinence des variables observées Les statistiques descriptives calculées pour chaque région permettent également d’avoir une vision globale de l’expression des différents traits dentaires (tables 6 à 10). Pour chaque aire étudiée nous présentons un tableau synthétisant l’adéquation entre les expressions des variables et leur utilité dans les futures analyses (figures 46 à 50). Chaque tableau est une reproduction des fiches d’enregistrement manuel des deux dentitions pour lesquelles nous avons mis en code des dégradés de gris permettant d’avoir directement les informations concernant l’état de leur expression. Nous avons dans un premier temps retiré (case cochée en gris foncé) les traits qui ont été éliminés lors du test de la concordance d’observation des variables. Nous avons ensuite mis en évidence les traits (case cochée en gris clair) qui présentent une expression constante, qui n’ont pas été observés ou lorsque l’effectif est trop faible, voire inexistant, pour être intégré même dans des phases de statistiques descriptives. Puis nous avons ajouté une étoile à une partie de ces dernières variables, celles que nous avons décidé d’éliminer de la suite de l’étude. Il s’agit avant tout de variables n’ayant jamais été observées ou à effectif réduit. Pour les traits qui expriment une expression constante, en revanche, même s’ils ne sont pas utiles lors des analyses régionales, ils peuvent se révéler l’être dans la synthèse de toutes les régions, c’est pourquoi nous ne les éliminons pas ici. Finalement, nous avons ajouté un tableau récapitulatif dans le coin supérieur gauche présentant, par dentition et par arcade, le nombre de traits de départ, le nombre de variables éliminées, la fréquence de cette valeur et le nombre de caractères restants. Une remarque s’impose quant à l’entoconulide (T56) qui est la 6e cuspide observée sur les molaires inférieures. Cette variable est enregistrée uniquement si la 5e cuspide (l’hypoconulide - T55) et la 6e cuspide sont présentes car si seule l’une d’entre elles est visible, il est impossible de déterminer laquelle. On rapporte alors les tailles respectives des deux cuspides. Les différents effectifs sont petits pour cette variable, mais nous décidons de ne par l’éliminer ici et réfléchirons à comment garder l’information qu’elle peut fournir dans les analyses, notamment par une éventuelle fusion des observations entre T55 et T56. 121 Chapitre 6 6.1.4.1. L’état des variables observées pour la Bohême Pour cette région, les données concernant les statistiques descriptives se trouvent en table 6. Le tableau cidessous indique qu’environ 35% (182) des variables sont soit à expression constante, soit à effectif réduit (figure 46). Pour la dentition permanente, cette valeur s’élève à environ 27% (92). Les traits observés sur la dentition inférieure sont en général moins variables avec 34% (49), les traits enregistrés sur le maxillaire quant à eux se situent à 23% (43). La dentition déciduale présente une valeur plus élevée à 48% (91). Comme pour la dentition permanente, les dents inférieures montrent une morphologie plus stable avec 63% (51) de traits constants, celles supérieures se situent vers 36% (40). Les caractères montrant le moins de variabilité sont, en général, ceux qui sont observés sur les racines. Nous choisissons d’éliminer uniquement les variables qui seront inexploitables lors des analyses. Il s’agit de traits observés sur la dentition déciduale, 36 pour le maxillaire et 16 pour la mandibule. Nb. traits Nb. éliminés DENTS PERMANENTES Droit Gauche Maxillaire M3 M2 M1 P2 P1 C I2 I1 I1 I2 C P1 P2 M1 M2 M3 1 Absence congénitale 2 Nombre de radicules 3 Racine appendiciforme 4 Nombre de racines des prémolaires 5 Nombre de racines des molaires 6 Racine de Carabelli 7 Racine paramolaire 8 Racine idiopathique 9 Degré de rotation 10 Convexité vestibulaire 11 Sillon traversant 12 Tubercule distal 13 Cingulum vestibulaire 14 Gouttière vestibulaire 15 Crêtes linguales marginales 16 Tubercule dentaire 17 Crêtes vestibulaires marginales 18 Crête mésiale 19 Crête accessoire distale 20 Cuspides accessoires 21 Odontome 22 Extension de l'émail 23 Parastyle 24 Trait de Carabelli 25 Métacone 26 Hypocone 27 Métaconule 28 Tubercule distal accessoire 29 Tubercule mésial paracone 30 Tubercule mésial accessoire 31 Protoconule 32 Tubercule lingual paracone Mandibule 33 Absence congénitale 34 Nombre de radicules 35 Racine appendiciforme 36 Nombre racines canines 37 Racine de Tomes 38 Nombre racines molaires 39 Racine de Citroen 40 Racine paramolaire 41 Racine entomolaire 42 Racine idiopathique 43 Crêtes linguales marginales 44 Crêtes vestibulaires marginales 45 Crête accessoire distale 46 Odontome 47 Variation des cuspides linguales 48 Pli cuspide mésiolinguale 49 Fosse précuspidale 50 Crête trigonide intermédiaire 51 Crête trigonide distale 52 Nombre de cuspides 53 Dessin du sillon 54 Protostylide 55 Hypoconulide 56 Entoconulide 57 Métaconulide 58 Tubercule paracone 59 Tubercule de Citroen Gauche M3 M2 M1 P2 P1 Droite C I2 I1 I1 I2 C P1 P2 M1 M2 M3 % Traits restants Permanentes 334 0 0% 334 Maxillaire Mandibule 186 148 0 0 0% 0% 186 148 Déciduales 186 52 27% 134 Maxillaire Mandibule 106 80 36 16 33% 20% 70 64 Dentition 520 468 DENTS DECIDUALES Droit Gauche Maxillaire m2 m1 c i2 i1 i1 i2 c m1 m2 1 Nombre de radicules 2 Extension de l'émail 3 Dilacération 4 Racine appendiciforme 5 Nombre de racines 6 Racine de la canine 7 Nombre de racines molaires 8 Racine de Carabelli 9 Racine paramolaire 10 Racine idiopathique 11 Forme des incisives 12 Gouttière vestibulaire 13 Crêtes linguales marginales 14 Tubercule dentaire 15 Déveleppoment cuspides 16 Crête du métacone 17 Parastyle 18 Trait de Carabelli 19 Métacone 20 Hypocone 21 Métaconule 22 Tubercule distal accessoire 23 Tubercule mésial paracone 24 Tubercule mésial accessoire 25 Protoconule 26 Tubercule lingual paracone Gauche Droite Mandibule m2 m1 c i2 i1 i1 i2 c m1 m2 27 Nombre de radicules 28 Extension de l'émail 29 Racine appendiciforme 30 Nombre de racines 31 Nombre de racines molaires 32 Racine de Citröen 33 Racine paramolaire 34 Racine idiopathique 35 Racine entomolaire 36 Forme des incisives 37 Double tubercule lingual 38 Développement cuspides 39 Protostylide 40 Fosse précuspidale 41 Distostylide fissuré 42 Crête trigonide intermédiaire 43 Crête trigonide distale 44 Pli mésiolingual 45 Hypoconulide 46 Entoconulide 47 Métaconulide 48 Tubercule paracone 49 Tubercule de Citröen 50 Fosse postérieure 51 Sillon trigonide 52 Proéminence cervicale Figure 46 : Représentation synthétique traduisant l’état des données pour la Bohême. En gris foncé, les traits éliminés lors du test de la concordance des modes d’observations des traits non métriques. En gris clair uniquement, les variables à expression constante et avec une étoile, les traits à observation insuffisante ou inexistante. 122 . Le traitement préliminaire des données 6.1.4.2. L’état des variables observées pour la Hongrie Pour la Hongrie, les données concernant les statistiques descriptives se trouvent en table 7. Le tableau cidessous indique qu’une large majorité des variables, environ 68% (356), sont soit à expression constante, soit à effectif réduit (figure 47). Pour la dentition permanente, cette valeur s’élève à environ 52% (171). En revanche, les traits qui ont été enregistrés sur la dentition déciduale ne possèdent en aucun cas un effectif suffisant (< 5) pour être intégrés dans l’étude. Les caractères montrant le moins de variabilité se répartissent sur l’ensemble de la dentition, même si la plupart d’entre eux s’observent sur les racines. Nous choisissons d’éliminer uniquement les variables qui seront inexploitables lors des analyses. Il s’agit ici des traits observés sur l’ensemble de la dentition déciduale. Nb. traits Nb. éliminés DENTS PERMANENTES Maxillaire Droit M3 M2 M1 P2 P1 Gauche C I2 I1 I1 I2 C P1 P2 M1 M2 M3 1 Absence congénitale 2 Nombre de radicules 3 Racine appendiciforme 4 Nombre de racines des prémolaires 5 Nombre de racines des molaires 6 Racine de Carabelli 7 Racine paramolaire 8 Racine idiopathique 9 Degré de rotation 10 Convexité vestibulaire 11 Sillon traversant 12 Tubercule distal 13 Cingulum vestibulaire 14 Gouttière vestibulaire 15 Crêtes linguales marginales 16 Tubercule dentaire 17 Crêtes vestibulaires marginales 18 Crête mésiale 19 Crête accessoire distale 20 Cuspides accessoires 21 Odontome 22 Extension de l'émail 23 Parastyle 24 Trait de Carabelli 25 Métacone 26 Hypocone 27 Métaconule 28 Tubercule distal accessoire 29 Tubercule mésial paracone 30 Tubercule mésial accessoire 31 Protoconule 32 Tubercule lingual paracone Mandibule 33 Absence congénitale 34 Nombre de radicules 35 Racine appendiciforme 36 Nombre racines canines 37 Racine de Tomes 38 Nombre racines molaires 39 Racine de Citroen 40 Racine paramolaire 41 Racine entomolaire 42 Racine idiopathique 43 Crêtes linguales marginales 44 Crêtes vestibulaires marginales 45 Crête accessoire distale 46 Odontome 47 Variation des cuspides linguales 48 Pli cuspide mésiolinguale 49 Fosse précuspidale 50 Crête trigonide intermédiaire 51 Crête trigonide distale 52 Nombre de cuspides 53 Dessin du sillon 54 Protostylide 55 Hypoconulide 56 Entoconulide 57 Métaconulide 58 Tubercule paracone 59 Tubercule de Citroen % Traits restants Permanentes 334 0 0% 334 Maxillaire Mandibule 186 148 0 0 0% 0% 186 148 Déciduales 186 186 100% 0 Maxillaire Mandibule 106 80 110 86 100% 100% 0 0 Dentition 520 DENTS DECIDUALES Maxillaire Gauche M3 M2 M1 P2 P1 Droite C I2 I1 I1 I2 C 334 Droit m2 m1 c Gauche i2 i1 i1 i2 i1 i1 i2 c m1 m2 1 Nombre de radicules 2 Extension de l'émail 3 Dilacération 4 Racine appendiciforme 5 Nombre de racines 6 Racine de la canine 7 Nombre de racines molaires 8 Racine de Carabelli 9 Racine paramolaire 10 Racine idiopathique 11 Forme des incisives 12 Gouttière vestibulaire 13 Crêtes linguales marginales 14 Tubercule dentaire 15 Déveleppoment cuspides 16 Crête du métacone 17 Parastyle 18 Trait de Carabelli 19 Métacone 20 Hypocone 21 Métaconule 22 Tubercule distal accessoire 23 Tubercule mésial paracone 24 Tubercule mésial accessoire 25 Protoconule 26 Tubercule lingual paracone P1 P2 M1 M2 M3 Mandibule Gauche m2 m1 c i2 Droite c m1 m2 27 Nombre de radicules 28 Extension de l'émail 29 Racine appendiciforme 30 Nombre de racines 31 Nombre de racines molaires 32 Racine de Citroen 33 Racine paramolaire 34 Racine idiopathique 35 Racine entomolaire 36 Forme des incisives 37 Double tubercule lingual 38 Développement cuspides 39 Protostylide 40 Fosse précuspidale 41 Distostylide fissuré 42 Crête trigonide intermédiaire 43 Crête trigonide distale 44 Pli mésiolingual 45 Hypoconulide 46 Entoconulide 47 Métaconulide 48 Tubercule paracone 49 Tubercule de Citroen 50 Fosse postérieure 51 Sillon trigonide 52 Proéminence cervicale Figure 47 : Représentation synthétique traduisant l’état des données pour la Hongrie. En gris foncé, les traits éliminés lors du test de la concordance des modes d’observations des traits non métriques. En gris clair uniquement, les variables à expression constante et avec une étoile, les traits à observation insuffisante ou inexistante. . 123 Chapitre 6 6.1.4.3. L’état des variables observées pour le sud de la France Pour cette région, les données concernant les statistiques descriptives se trouvent en table 8. Le tableau cidessous indique qu’environ 37% (189) des variables sont soit à expression constante, soit à effectif réduit (figure 48). On trouve ici des valeurs comparables à celles de la Bohême. Pour la dentition permanente, cette valeur s’élève à environ 26% (87). Les traits observés sur la dentition inférieure sont en général moins variables avec 33% (47), les traits enregistrés sur le maxillaire se situent, quant à eux, à 22% (40). La dentition déciduale présente une valeur plus élevée à 62% (115). Les dents inférieures montrent une morphologie plus stable avec 73% (59) de traits constants, celles inférieures se situent vers 53% (56). Les caractères montrant le moins de variabilité sont, en général, ceux observés sur les racines. Nous gardons toutes les variables, la dentition déciduale possédant des effectifs qui pourraient, selon les traits, être exploitables lors des analyses. Nb. traits Nb. éliminés DENTS PERMANENTES Maxillaire Droit M3 M2 M1 P2 P1 Gauche C I2 I1 I1 I2 C P1 P2 M1 M2 M3 1 Absence congénitale 2 Nombre de radicules 3 Racine appendiciforme 4 Nombre de racines des prémolaires 5 Nombre de racines des molaires 6 Racine de Carabelli 7 Racine paramolaire 8 Racine idiopathique 9 Degré de rotation 10 Convexité vestibulaire 11 Sillon traversant 12 Tubercule distal 13 Cingulum vestibulaire 14 Gouttière vestibulaire 15 Crêtes linguales marginales 16 Tubercule dentaire 17 Crêtes vestibulaires marginales 18 Crête mésiale 19 Crête accessoire distale 20 Cuspides accessoires 21 Odontome 22 Extension de l'émail 23 Parastyle 24 Trait de Carabelli 25 Métacone 26 Hypocone 27 Métaconule 28 Tubercule distal accessoire 29 Tubercule mésial paracone 30 Tubercule mésial accessoire 31 Protoconule 32 Tubercule lingual paracone Mandibule 33 Absence congénitale 34 Nombre de radicules 35 Racine appendiciforme 36 Nombre racines canines 37 Racine de Tomes 38 Nombre racines molaires 39 Racine de Citroen 40 Racine paramolaire 41 Racine entomolaire 42 Racine idiopathique 43 Crêtes linguales marginales 44 Crêtes vestibulaires marginales 45 Crête accessoire distale 46 Odontome 47 Variation des cuspides linguales 48 Pli cuspide mésiolinguale 49 Fosse précuspidale 50 Crête trigonide intermédiaire 51 Crête trigonide distale 52 Nombre de cuspides 53 Dessin du sillon 54 Protostylide 55 Hypoconulide 56 Entoconulide 57 Métaconulide 58 Tubercule paracone 59 Tubercule de Citroen % Traits restants Permanentes 334 0 0% 334 Maxillaire Mandibule 186 148 0 0 0% 0% 186 148 Déciduales 186 0 0% 186 Maxillaire Mandibule 106 80 0 0 0% 0% 106 80 Dentition 520 DENTS DECIDUALES Maxillaire Gauche M3 M2 M1 P2 P1 Droite C I2 I1 I1 I2 C 520 Droit m2 m1 c Gauche i2 i1 i1 i2 i1 i1 i2 c m1 m2 1 Nombre de radicules 2 Extension de l'émail 3 Dilacération 4 Racine appendiciforme 5 Nombre de racines 6 Racine de la canine 7 Nombre de racines molaires 8 Racine de Carabelli 9 Racine paramolaire 10 Racine idiopathique 11 Forme des incisives 12 Gouttière vestibulaire 13 Crêtes linguales marginales 14 Tubercule dentaire 15 Déveleppoment cuspides 16 Crête du métacone 17 Parastyle 18 Trait de Carabelli 19 Métacone 20 Hypocone 21 Métaconule 22 Tubercule distal accessoire 23 Tubercule mésial paracone 24 Tubercule mésial accessoire 25 Protoconule 26 Tubercule lingual paracone P1 P2 M1 M2 M3 Mandibule Gauche m2 m1 c i2 Droite c m1 m2 27 Nombre de radicules 28 Extension de l'émail 29 Racine appendiciforme 30 Nombre de racines 31 Nombre de racines molaires 32 Racine de Citroen 33 Racine paramolaire 34 Racine idiopathique 35 Racine entomolaire 36 Forme des incisives 37 Double tubercule lingual 38 Développement cuspides 39 Protostylide 40 Fosse précuspidale 41 Distostylide fissuré 42 Crête trigonide intermédiaire 43 Crête trigonide distale 44 Pli mésiolingual 45 Hypoconulide 46 Entoconulide 47 Métaconulide 48 Tubercule paracone 49 Tubercule de Citroen 50 Fosse postérieure 51 Sillon trigonide 52 Proéminence cervicale Figure 48 : Représentation synthétique traduisant l’état des données pour le sud de la France. En gris foncé, les traits éliminés lors du test de la concordance des modes d’observations des traits non métriques. En gris clair uniquement, les variables à expression constante. 124 . Le traitement préliminaire des données 6.1.4.4. L’état des variables observées pour le nord de l’Espagne Pour cette région, les données concernant les statistiques descriptives se trouvent en table 9. Le tableau cidessous indique qu’une majorité des variables, environ 65% (337), sont soit à expression constante, soit à effectif réduit (figure 49). Pour la dentition permanente, cette valeur s’élève à environ 45% (151). En revanche, les traits qui ont été enregistrés sur la dentition déciduale ne possèdent en aucun cas un effectif suffisant pour être intégrés dans l’étude. Les caractères montrant le moins de variabilité se répartissent sur l’ensemble de la dentition, même si la plupart d’entre eux s’observent sur les racines. Nous choisissons d’éliminer les variables qui seront inexploitables lors des analyses. Il s’agit ici des traits observés sur l’ensemble de la dentition déciduale. Nb. traits Nb. éliminés DENTS PERMANENTES Maxillaire Droit M3 M2 M1 P2 P1 Gauche C I2 I1 I1 I2 C P1 P2 M1 M2 M3 1 Absence congénitale 2 Nombre de radicules 3 Racine appendiciforme 4 Nombre de racines des prémolaires 5 Nombre de racines des molaires 6 Racine de Carabelli 7 Racine paramolaire 8 Racine idiopathique 9 Degré de rotation 10 Convexité vestibulaire 11 Sillon traversant 12 Tubercule distal 13 Cingulum vestibulaire 14 Gouttière vestibulaire 15 Crêtes linguales marginales 16 Tubercule dentaire 17 Crêtes vestibulaires marginales 18 Crête mésiale 19 Crête accessoire distale 20 Cuspides accessoires 21 Odontome 22 Extension de l'émail 23 Parastyle 24 Trait de Carabelli 25 Métacone 26 Hypocone 27 Métaconule 28 Tubercule distal accessoire 29 Tubercule mésial paracone 30 Tubercule mésial accessoire 31 Protoconule 32 Tubercule lingual paracone Mandibule 33 Absence congénitale 34 Nombre de radicules 35 Racine appendiciforme 36 Nombre racines canines 37 Racine de Tomes 38 Nombre racines molaires 39 Racine de Citroen 40 Racine paramolaire 41 Racine entomolaire 42 Racine idiopathique 43 Crêtes linguales marginales 44 Crêtes vestibulaires marginales 45 Crête accessoire distale 46 Odontome 47 Variation des cuspides linguales 48 Pli cuspide mésiolinguale 49 Fosse précuspidale 50 Crête trigonide intermédiaire 51 Crête trigonide distale 52 Nombre de cuspides 53 Dessin du sillon 54 Protostylide 55 Hypoconulide 56 Entoconulide 57 Métaconulide 58 Tubercule paracone 59 Tubercule de Citroen % Traits restants Permanentes 334 0 0% 334 Maxillaire Mandibule 186 148 0 0 0% 0% 186 148 Déciduales 186 186 100% 0 Maxillaire Mandibule 106 80 110 86 100% 100% 0 0 Dentition 520 DENTS DECIDUALES Maxillaire Gauche M3 M2 M1 P2 P1 Droite C I2 I1 I1 I2 C 334 Droit m2 m1 c Gauche i2 i1 i1 i2 i1 i1 i2 c m1 m2 1 Nombre de radicules 2 Extension de l'émail 3 Dilacération 4 Racine appendiciforme 5 Nombre de racines 6 Racine de la canine 7 Nombre de racines molaires 8 Racine de Carabelli 9 Racine paramolaire 10 Racine idiopathique 11 Forme des incisives 12 Gouttière vestibulaire 13 Crêtes linguales marginales 14 Tubercule dentaire 15 Déveleppoment cuspides 16 Crête du métacone 17 Parastyle 18 Trait de Carabelli 19 Métacone 20 Hypocone 21 Métaconule 22 Tubercule distal accessoire 23 Tubercule mésial paracone 24 Tubercule mésial accessoire 25 Protoconule 26 Tubercule lingual paracone P1 P2 M1 M2 M3 Mandibule Gauche m2 m1 c i2 Droite c m1 m2 27 Nombre de radicules 28 Extension de l'émail 29 Racine appendiciforme 30 Nombre de racines 31 Nombre de racines molaires 32 Racine de Citroen 33 Racine paramolaire 34 Racine idiopathique 35 Racine entomolaire 36 Forme des incisives 37 Double tubercule lingual 38 Développement cuspides 39 Protostylide 40 Fosse précuspidale 41 Distostylide fissuré 42 Crête trigonide intermédiaire 43 Crête trigonide distale 44 Pli mésiolingual 45 Hypoconulide 46 Entoconulide 47 Métaconulide 48 Tubercule paracone 49 Tubercule de Citroen 50 Fosse postérieure 51 Sillon trigonide 52 Proéminence cervicale Figure 49 : Représentation synthétique traduisant l’état des données pour le nord de l’Espagne. En gris foncé, les traits éliminés lors du test de la concordance des modes d’observations des traits non métriques. En gris clair uniquement, les variables à expression constante et avec une étoile, les traits à observation insuffisante ou inexistante. . 125 Chapitre 6 6.1.4.5. L’état des variables observées pour la Suisse La Suisse est la région qui montre la plus grande variabilité d’expression des traits dentaires. Les données concernant les statistiques descriptives se trouvent en table 10. Le tableau ci-dessous indique que seul 28% (148) des variables sont soit à expression constante, soit à effectif réduit (figure 50). Pour la dentition permanente, cette valeur s’élève à environ 20% (67). Les traits observés sur la dentition inférieure sont en général moins variables avec 26% (38), les traits enregistrés sur le maxillaire quant à eux se situent à 16% (29). La dentition déciduale présente une valeur plus élevée à 44% (81). Comme pour la dentition permanente, les dents inférieures montrent une morphologie plus stable avec 51% (41) de traits constants, celles qui sont supérieures se situent vers 38% (40). Les caractères montrant le moins de variabilité sont, en général, ceux observés sur les racines. Nous gardons toutes les variables, la dentition déciduale possédant des effectifs qui pourraient selon les traits être exploitables lors des analyses. Nb. traits Nb. éliminés DENTS PERMANENTES Maxillaire Droit M3 M2 M1 P2 P1 Gauche C I2 I1 I1 I2 C P1 P2 M1 M2 M3 1 Absence congénitale 2 Nombre de radicules 3 Racine appendiciforme 4 Nombre de racines des prémolaires 5 Nombre de racines des molaires 6 Racine de Carabelli 7 Racine paramolaire 8 Racine idiopathique 9 Degré de rotation 10 Convexité vestibulaire 11 Sillon traversant 12 Tubercule distal 13 Cingulum vestibulaire 14 Gouttière vestibulaire 15 Crêtes linguales marginales 16 Tubercule dentaire 17 Crêtes vestibulaires marginales 18 Crête mésiale 19 Crête accessoire distale 20 Cuspides accessoires 21 Odontome 22 Extension de l'émail 23 Parastyle 24 Trait de Carabelli 25 Métacone 26 Hypocone 27 Métaconule 28 Tubercule distal accessoire 29 Tubercule mésial paracone 30 Tubercule mésial accessoire 31 Protoconule 32 Tubercule lingual paracone Mandibule 33 Absence congénitale 34 Nombre de radicules 35 Racine appendiciforme 36 Nombre racines canines 37 Racine de Tomes 38 Nombre racines molaires 39 Racine de Citroen 40 Racine paramolaire 41 Racine entomolaire 42 Racine idiopathique 43 Crêtes linguales marginales 44 Crêtes vestibulaires marginales 45 Crête accessoire distale 46 Odontome 47 Variation des cuspides linguales 48 Pli cuspide mésiolinguale 49 Fosse précuspidale 50 Crête trigonide intermédiaire 51 Crête trigonide distale 52 Nombre de cuspides 53 Dessin du sillon 54 Protostylide 55 Hypoconulide 56 Entoconulide 57 Métaconulide 58 Tubercule paracone 59 Tubercule de Citroen % Traits restants Permanentes 334 0 0% 334 Maxillaire Mandibule 186 148 0 0 0% 0% 186 148 Déciduales 186 0 0% 186 Maxillaire Mandibule 106 80 0 0 0% 0% 106 80 Dentition 520 DENTS DECIDUALES Maxillaire Gauche M3 M2 M1 P2 P1 Droite C I2 I1 I1 I2 C 520 Droit m2 m1 c Gauche i2 i1 i1 i2 i1 i1 i2 c m1 m2 1 Nombre de radicules 2 Extension de l'émail 3 Dilacération 4 Racine appendiciforme 5 Nombre de racines 6 Racine de la canine 7 Nombre de racines molaires 8 Racine de Carabelli 9 Racine paramolaire 10 Racine idiopathique 11 Forme des incisives 12 Gouttière vestibulaire 13 Crêtes linguales marginales 14 Tubercule dentaire 15 Déveleppoment cuspides 16 Crête du métacone 17 Parastyle 18 Trait de Carabelli 19 Métacone 20 Hypocone 21 Métaconule 22 Tubercule distal accessoire 23 Tubercule mésial paracone 24 Tubercule mésial accessoire 25 Protoconule 26 Tubercule lingual paracone P1 P2 M1 M2 M3 Mandibule Gauche m2 m1 c i2 Droite c m1 m2 27 Nombre de radicules 28 Extension de l'émail 29 Racine appendiciforme 30 Nombre de racines 31 Nombre de racines molaires 32 Racine de Citroen 33 Racine paramolaire 34 Racine idiopathique 35 Racine entomolaire 36 Forme des incisives 37 Double tubercule lingual 38 Développement cuspides 39 Protostylide 40 Fosse précuspidale 41 Distostylide fissuré 42 Crête trigonide intermédiaire 43 Crête trigonide distale 44 Pli mésiolingual 45 Hypoconulide 46 Entoconulide 47 Métaconulide 48 Tubercule paracone 49 Tubercule de Citroen 50 Fosse postérieure 51 Sillon trigonide 52 Proéminence cervicale Figure 50 : Représentation synthétique traduisant l’état des données pour la Suisse. En gris foncé, les traits éliminés lors du test de la concordance des modes d’observations des traits non métriques. En gris clair uniquement, les variables à expression constante. 126 . Le traitement préliminaire des données 6.1.4.6. L’état des variables observées : un bilan Selon les régions étudiées, l’état des variables peut être relativement différent. La composition des échantillons en est en grande partie responsable. Nous nous sommes intéressée non seulement aux traits qui ne seront pas mobilisables pour la suite des analyses, mais également à ceux présentant une expression constante. • L’éviction des traits non mobilisables Nous avons choisi d’éliminer les variables qui n’apporteraient aucune information utile lors du traitement préliminaire des données de par leur nombre de données restreint ou simplement parce qu’elles n’ont jamais été observées. Les traits à expression constante ne sont pas supprimés ici. De ce fait, seules deux régions (la Bohême et la Hongrie) ont fait l’objet de l’élimination de variables, observées uniquement sur la dentition déciduale. L’effectif insuffisant est la raison principale de leur suppression. Nous avons fait abstraction des données pour l’entoconulide (T56) pour les raisons évoquées au préalable. Nous nous pencherons sur le sort de cette variable plus loin. • Les traits à expression constante Les représentations schématiques (figures 46 à 50) révèlent des tendances similaires pour les différents échantillons observés. Les variables à expression constante sont de deux types. Celles unilatéralement invariantes (seul l’un des deux antimères est à expression constante) et peuvent ainsi être encore mobilisés pour les analyses. Celles bilatéralement invariantes (les deux antimères sont à expressions constantes) et seront pertinentes uniquement, mais non obligatoirement, lors des analyses comprenant l’ensemble des régions. Nous n’avons pas éliminé ici ce deuxième type de variables, même si le traitement préliminaire des données n’est pas nécessaire. Les différents tests n’en tiendront tout simplement pas compte. La dentition permanente Selon les échantillons, les variables à expression constante diffèrent, cependant les mêmes tendances se dessinent, avec : - une grande majorité de traits à expression invariante appartenant au système FU-DTS (Freiburg University - dental traits system). Ce système propose des variables rarement exprimées, et à l’évidence, cette situation se reflète bien ici - une partie des variables constantes sont celles observées sur les racines. Nous ne sommes pas réellement surprise puisque cette partie de la dent est celle qui montre le moins de variabilité - les dents inférieures et plus particulièrement celles antérieures (incisives et canines) sont les moins variables La dentition déciduale D’une manière générale, les dents lactéales montrent très peu de variabilité morphologique. De ce fait, la majorité des traits à expression constante sont localisés sur cette dentition. Il est tout de même possible de suivre les mêmes tendances observées sur la dentition permanente, avec : - un pourcentage élevé de traits appartenant au système FU-DTS - une large quantité de variables observées sur les racines - des dents inférieures nettement moins variables que les supérieures, et cette remarque s’impose d’autant plus ici, puisque les incisives et les canines sont d’une constance imperturbable 127 Chapitre 6 6.2. L’asymétrie des antimères Cette étape du traitement préliminaire des données est la plus importante et la plus utile puisqu’elle vise avant tout à obtenir des variables unilatérales en gardant le meilleur reflet de l’apport des gènes dans l’expression des différents traits. De plus, elle permet l’élimination de données redondantes en réduisant les données manquantes. S’il est vrai qu’au premier abord les dents controlatérales semblent identiques, observées de plus près, les variations morphologiques montrent parfois une asymétrie qui se traduit soit par une différence dans le degré d’expression, soit par la présence ou l’absence d’un trait (Biggerstaff 1979). Les différentes structures bilatérales du corps humain disposent des mêmes instructions génétiques de départ. L’asymétrie qu’elles sont susceptibles de présenter est due à des interférences se produisant pendant le développement (Bailit et al. 1970). C.H. Waddington (1957) introduit la notion de canalisation du développement (buffering). Il s’agit de la disposition d’un individu à se protéger contre un stress environnemental. Ainsi, selon leur intensité, les influences environnementales et la capacité de canalisation du développement peuvent dévier un phénotype de son génotype de manière plus ou moins importante (Townsend et Martin 1992). Ce concept permet de comprendre, en partie, la présence d’asymétrie sur les antimères (Naugler et Ludman 1996). L. Van Valen (1962) a mis en évidence trois types primaires d’asymétrie qui peuvent être présents sur les structures bilatérales (figure 51). Celle qui caractérise les traits non métriques dentaires est l’asymétrie fluctuante, plus communément appelée dans la littérature random asymmetry - asymétrie aléatoire -. L’asymétrie directionnelle Terme employé pour décrire des structures bilatérales dont un côté montre un développement régulier plus fort que l’autre. L’antisymétrie Terme employé pour décrire des structures qui sont en général asymétriques et qui ne montrent pas de côté préférentiel dans leur asymétrie. L’asymétrie fluctuante Terme employé pour décrire des structures qui sont normalement symétriques mais qui peuvent montrer des différences sans côté préférentiel. Figure 51 : Les différents types d’asymétrie liés à des interférences développementales définis par L. Van Valen (1962). . Comme toutes les structures bilatérales, les dents ont fait l’objet de centaines d’études ayant pour objectif de cerner les causes et l’intensité de l’asymétrie. Malgré une littérature abondante, les recherches ont principalement touché les données métriques dentaires (Potter et Nance 1976, Kolakowski et Bailit 1981, Guagliardo 1982, Smith et al. 1982, Boklage 1987, Kieser et Groeneveld 1988, Harila 2004...). Nous pourrions imaginer nous servir des résultats concernant les données métriques pour tenter d’expliquer les raisons de l’asymétrie sur les traits morphologiques, mais les relations entre l’asymétrie des variables métriques et non métriques dentaires ne sont pas encore réellement déterminées. Selon les études, il n’y a pas de corrélation entre l’asymétrie de ces deux types de variables (Noss et al. 1983, Saunders et Mayhall 1986), ou alors la corrélation se révèle négative (Baume et Crawford 1980). Un certain nombre d’études sur les traits non métriques dentaires ont tout de même essayé de caractériser et de préciser les raisons de la présence de l’asymétrie sur ce type de variables. Certains auteurs ont recherché si l’asymétrie des antimères pouvait être héréditaire. R.N. Staley et L.R. Green (1974) ont travaillé à la fois sur 180 paires de jumeaux (monozygotes et dizygotes) et sur un échantillon de plus de 1000 individus nord-américains. D’après leurs résultats, l’asymétrie entre les dents controlatérales ne semble pas être héréditaire. D’autres études (Greene 1982, Mizoguchi 1989, Staley et al. 1998...), réalisées sur des jumeaux ou sur des individus apparentés, sont allées dans le même sens. 128 Le traitement préliminaire des données Les recherches ont également abordé la question de l’influence du sexe des individus sur la présence et l’intensité de l’asymétrie. Selon les études, les résultats peuvent être contradictoires. Les travaux de R.N. Staley et L.R. Green (1974), A.J. Perzigian (1976) ou encore J.F. Noss et al. (1983) n’ont mis en évidence aucun dimorphisme sexuel dans l'expression de l’asymétrie des traits non métriques dentaires. En revanche, Y. Mizoguchi (1988) a enregistré une fréquence plus élevée de l’asymétrie pour la femme dans certaines populations asiatiques, fréquence qui ne s’est pas révélée statistiquement significative. La plupart des recherches ont toutefois montré que les traits dentaires exprimés sur les dents controlatérales présentent en général un fort degré de symétrie. Les concordances de présence et d’absence des variables s’élèvent à plus de 90%. Ces résultats se situent entre 50 et 80% si toutes les formes d’expressions sont prises en compte (Saunders et Mayhall 1982). Néanmoins, certains travaux, comme celui de R.H. Biggerstaff (1973) notamment, montrent que les niveaux de symétrie peuvent parfois atteindre des taux nettement inférieurs. Les tenants et aboutissants de l’asymétrie dentaire ne sont pas déterminés avec précision. Pour résoudre le problème lié à l’asymétrie des expressions des variations morphologiques sur les dents controlatérales, plusieurs moyens d’enregistrement des données sont utilisés. Certains conservent l’enregistrement bilatéral des différentes variables, cependant ce procédé est discutable puisqu’il augmente l’effectif des échantillons (Mayhall 1992). Une seule observation pour chaque caractère est nécessaire pour ne pas augmenter démesurément la taille du corpus (Nichol 1989). On peut donc se contenter de l’enregistrement unilatéral pour éviter ce problème, mais cela réduit la taille de l’échantillon. Chaque méthode a ses inconvénients, c’est pourquoi certains auteurs, comme G.R. Scott et C.G. Turner (Scott 1977, Turner et Scott 1977), recommandent d’observer la dentition complète, ensuite seule l’expression la plus élevée observée sur les antimères est maintenue. Si l’expression d’une variable sur les antimères est asymétrique, on considère la plus développée comme le meilleur reflet de l’apport des gènes (Scott et Turner 1997). En ce qui concerne cette étude, nous avons enregistré les variables sur l’ensemble de la dentition. C’est pourquoi, nous procéderons, dans un premier temps, à une phase de vérification de la similarité de l’expression des traits sur les antimères par une mesure de corrélation. Dans un second temps, nous nous attacherons aux modalités du traitement des antimères selon les résultats de leur degré de similarité, le type de variables et l’état des différents échantillons. 6.2.1. Le degré de similarité des antimères Selon les régions, la composition des échantillons est très différente (dents isolées, dents sur arcade, situation mixte). Les corpus pour la Bohême et la Hongrie se présentent sous forme d’individus où la mesure du degré de similarité des antimères peut être réalisée. En revanche, le sud de la France et le nord de l’Espagne sont constitués, pour la plupart, de tombes collectives et, par conséquent, par un nombre relativement important de dents isolées. Les calculs ne seront pas des plus aisés puisqu’ils se baseront sur un nombre restreint d’antimères. La Suisse, quant à elle, présente une situation intermédiaire. Ainsi, selon les échantillons, une prudence est nécessaire concernant les interprétations que nous pourrions inférer à partir des résultats. Nous souhaitons mettre en évidence les relations existantes entre les expressions des variables sur les dents controlatérales. Nous l’effectuons par une mesure non paramétrique d’association pour variables ordinales ou graduées, le Tau de Kendall. Nous déterminons les hypothèses de départ suivantes : H0 = hypothèse nulle : les expressions sur les dents controlatérales sont interdépendantes, aucune asymétrie significative ne peut être mise en évidence. H1 = hypothèse alternative : les expressions sur les dents controlatérales ne sont pas interdépendantes, le niveau d’asymétrie est trop important pour qu’il soit purement aléatoire. Les hypothèses sont testées au seuil de signification de 0.05 ou 0.01. Selon le seuil de signification retenu, les expressions des antimères sont statistiquement corrélées si leur valeur est inférieure au seuil. Le calcul des corrélations s’effectue sur 259 paires de variables (518 variables unilatérales). 129 Chapitre 6 6.2.1.1. Le résultat des corrélations entre les antimères pour la Bohême L’échantillon composé de tombes individuelles, et donc par des sujets personnalisés, permet d’effectuer ce test sur un nombre important de variables (figure 52). Sur les 235 traits de départ (24 ayant été éliminés lors de phases précédentes), 131 caractères (58%) ont fait l’objet d’un calcul. Les résultats de cette mesure se trouvent en table 11. Il s’agit, plus précisément, de 104 (62%) variables pour la dentition permanente et 27 (40%) pour la déciduale. Sur l’ensemble de ces traits, seules la racine idiopathique observée sur la troisième molaire supérieure (T8) et l’extension de l’émail observée sur la deuxième prémolaire supérieure (T22) ne répondent pas à l’exigence de l’hypothèse nulle (H0). Par conséquent, 129 variables sur 131 rejettent l’hypothèse alternative (H1) et montrent que leurs expressions sur les dents controlatérales sont interdépendantes. Les causes pour lesquelles les mesures n’ont pas pu être réalisées sur certaines variables (104 traits, dont 63 observés sur la dentition permanente et 41 sur la déciduale) se répartissent de la manière suivante : 4 traits observés sur la dentition déciduale ne possèdent plus qu’un seul antimère à ce stade du traitement préliminaire, 63 traits disposent de deux antimères à expression constante, pour 24 autres variables un seul des deux antimères est à expression constante et pour les 13 derniers caractères les paires d’observations sont constantes. Figure 52 : Représentation synthétique touchant les résultats du degré de similarité de l’asymétrie des antimères pour la Bohême. En gris foncé, les variables éliminées lors des phases antérieures du traitement préliminaire des données. 130 Le traitement préliminaire des données 6.2.1.2. Le résultat des corrélations entre les antimères pour la Hongrie L’échantillon est constitué de sujets, pour la plupart, individualisés, ce qui permet d’effectuer ce test sur un nombre important de variables. L’état général du matériel dentaire étant ce qu’il est, les données sur les antimères ici reposent sur un nombre restreint d’individus et de variables (figure 53). Sur les 167 traits de départ (toutes les observations réalisées sur la dentition déciduale ayant été éliminées lors de la phase précédente), seuls 65 caractères (39%) ont fait l’objet d’un calcul. Les résultats de cette mesure se trouvent en table 12. Sur l’ensemble de ces traits, seuls le dessin du sillon (T53) et l’hypoconulide (T55), observés tous deux sur la troisième molaire inférieure, ne répondent pas à l’exigence de l’hypothèse nulle (H0). Et, par conséquent, 63 variables sur 65 rejettent l’hypothèse alternative (H1) et montrent que leurs expressions sur les dents controlatérales sont interdépendantes. Les causes pour lesquelles les mesures n’ont pas pu être réalisées sur certaines variables (102 traits) se répartissent de la manière suivante : 75 traits disposent de deux antimères à expression constante, pour 17 variables un seul des deux antimères est à expression constante et pour les 10 derniers caractères les paires d’observations sont constantes. Figure 53 : Représentation synthétique touchant les résultats du degré de similarité de l’asymétrie des antimères pour la Hongrie. En gris foncé, les variables éliminées lors des phases antérieures du traitement préliminaire des données. 131 Chapitre 6 6.2.1.3. Le résultat des corrélations entre les antimères pour le sud de la France Cette région est constituée essentiellement de dents isolées, c’est pourquoi, sur les 259 traits de départ, seuls 78 caractères (31%) ont fait l’objet d’un calcul (figure 54). Les résultats de cette mesure se trouvent en table 13. Il s’agit, plus précisément, de 65 (40%) variables pour la dentition permanente et 13 (14%) pour la déciduale. Sur l’ensemble de ces traits, six variables ne répondent pas à l’exigence de l’hypothèse nulle (H0). Il s’agit, pour la dentition permanente, du nombre de racines de la première molaire supérieure (T5), du sillon traversant de la seconde incisive supérieure (T11), du tubercule accessoire distal observé sur la première molaire supérieure (T28), du dessin du sillon des première et deuxième molaires inférieures (T53), et pour la dentition déciduale, des crêtes linguales marginales de la canine supérieure (T14). Par conséquent, 72 variables sur 78 rejettent l’hypothèse alternative (H1) montrant que leurs expressions sur les antimères sont interdépendantes. Les causes pour lesquelles les mesures n’ont pas pu être réalisées sur certaines variables (181 traits, dont 102 observés sur la dentition permanente et 79 sur la déciduale) se répartissent de la manière suivante : 83 traits disposent de deux antimères à expression constante, pour 39 autres variables un seul des deux antimères est à expression constante et pour les 59 derniers caractères les paires d’observations sont constantes. Figure 54 : Représentation synthétique touchant les résultats du degré de similarité de l’asymétrie des antimères pour le sud de la France. En gris foncé, les variables éliminées lors des phases antérieures du traitement préliminaire des données. 132 Le traitement préliminaire des données 6.2.1.4. Le résultat des corrélations entre les antimères pour le nord de l’Espagne L’échantillon est largement composé de dents isolées, c’est pourquoi, sur les 167 traits de départ (toutes les observations réalisées sur la dentition déciduale ayant été éliminées lors de la phase précédente), seuls 55 caractères (33%) ont fait l’objet d’un calcul (figure 55). Les résultats de cette mesure se trouvent en table 14. Sur l’ensemble de ces traits, huit traits ne répondent pas à l’exigence de l’hypothèse nulle (H0). Il s’agit, pour la dentition supérieure, du nombre de radicules de la troisième molaire (T2), du nombre de racines de la première prémolaire (T4) et de la troisième molaire (T5), du tubercule distal de l’incisive centrale (T12), et du trait de Carabelli pour la troisième molaire (T24) ; pour celle inférieure, du nombre de radicules de la première molaire (T34), du nombre de racines de la troisième molaire (T38) et du protostylide de la troisième molaire (T54). Et, par conséquent, 47 variables sur 55 rejettent l’hypothèse alternative (H1) et montrent que leurs expressions sur les dents controlatérales sont interdépendantes. Les causes pour lesquelles les mesures n’ont pas pu être réalisées sur certaines variables (112 traits) se répartissent de la manière suivante : 66 traits disposent de deux antimères à expression constante, pour 21 autres variables un seul des deux antimères est à expression constante et pour les 25 derniers caractères les paires d’observations sont constantes. Figure 55 : Représentation synthétique touchant les résultats du degré de similarité de l’asymétrie des antimères pour le nord de l’Espagne. En gris foncé, les variables éliminées lors des phases antérieures du traitement préliminaire des données. 133 Chapitre 6 6.2.1.5. Le résultat des corrélations entre les antimères pour la Suisse Les échantillons de la Suisse sont composés à la fois de dents isolées et de sujets individualisés. Sur les 259 traits de départ, 148 caractères (57%) ont fait l’objet d’un calcul (figure 56). Les résultats de cette mesure se trouvent en table 15. Il s’agit, plus précisément, de 105 (63%) variables pour la dentition permanente et 43 (47%) pour la déciduale. Sur l’ensemble de ces traits, seules trois variables observées sur la dentition permanente ne répondent pas à l’exigence de l’hypothèse nulle (H0). Il s’agit du tubercule lingual paracone examiné sur la première molaire supérieure (T32), du métaconulide observé sur la première molaire inférieure (T57) et du tubercule paracone enregistré sur la deuxième molaire inférieure (T58). Par conséquent, 102 variables sur 105 rejettent l’hypothèse alternative (H1) et montrent que leurs expressions sur les dents controlatérales sont interdépendantes. Les causes pour lesquelles les mesures n’ont pas pu être réalisées sur certaines variables (107 traits, dont 62 observés sur la dentition permanente et 49 sur la déciduale) se répartissent de la manière suivante : 59 traits disposent de deux antimères à expression constante, pour 28 autres variables un seul des deux antimères est à expression constante et pour les 24 derniers caractères les paires d’observations sont constantes. Figure 56 : Représentation synthétique touchant les résultats du degré de similarité de l’asymétrie des antimères pour la Suisse. En gris foncé, les variables éliminées lors des phases antérieures du traitement préliminaire des données. 134 Le traitement préliminaire des données 6.2.1.6. Le résultat des corrélations entre les antimères : un bilan Les différentes variables observées sur l’ensemble des régions étudiées ont fait l’objet d’une mesure du degré de similarité de leur expression sur les antimères. Le calcul de la ressemblance des variations morphologiques et de leur intensité sur les dents controlatérales est tributaire de la spécificité des échantillons (dents isolées ou sujets individualisés), et non de leur nombre. Ainsi, les données provenant des régions occidentales (nord de l’Espagne et sud de la France) ne sont pas celles qui ont fourni le plus d’informations. En effet, les échantillons qui les composent sont constitués dans la grande majorité de dents isolées et, sans surprise, peu de calculs ont pu être réalisés. En revanche, la Suisse présentant une situation mixte est la région qui a procuré le plus d’indications. Ainsi, selon le territoire considéré, le nombre de variables qui a pu être testé se situe entre 30 et 60%. D’une manière générale, lorsque la mesure a été réalisée, les résultats indiquent clairement que les dents controlatérales montrent un haut degré de symétrie (entre 86 et 98% avec une moyenne à plus de 94%) répondant ainsi à l’exigence de l’hypothèse nulle (HO) avec un seuil d’acceptation de 0.01 pour l’ensemble des variables (à l’exception de quelques traits présentant un seuil de 0.05, soit deux variables pour l’échantillon suisse et trois pour le nord de l’Espagne). Un nombre restreint de variables se sont dévoilées asymétriques dans leur expression bilatérale. En voici un récapitulatif : • T2-M3-nord de l’Espagne • T32-M1-Suisse • T4-P1-nord de l’Espagne • T34-M1-nord de l’Espagne • T5-M1-sud de la France/M3-nord de l’Espagne • T38-M3-nord de l’Espagne • T8-M3-Bohême • T53-M1-M2-sud de la France/M3-Hongrie • T11-I2-sud de la France • T54-M3-nord de l’Espagne • T12-I1-nord de l’Espagne • T55-M3-Hongrie • T22-P2-Bohême • T57-M1-Suisse • T24-M3-nord de l’Espagne • T58-M2-Suisse • T28-M1-sud de la France • T14-c-sud de la France Ces traits, à l’évidence, varient d’un échantillon à l’autre. Nous attribuons clairement la présence de ces antimères non corrélés à des biais d’échantillonnage. En effet, sur l’ensemble de ces variables (n=21), 14 d’entre elles proviennent d’échantillons composés en majorité de dents isolées (sud de la France et nord de l’Espagne) pour lesquels les calculs ont été effectués sur des effectifs restreints en observations bilatérales. De plus, nos recherches nous ont amenée à observer une quantité non négligeable de traits dentaires et nous savons qu’ils s’expriment très largement, voire unanimement, sur les dents controlatérales. Une exception peut néanmoins être relevée avec le sillon traversant (T11), trait observé sur les incisives supérieures. Lorsqu’un sillon est présent sur les antimères, parfois sa position diffère (mésiale, distale, médiane ou mésio-distale). Même si la non-corrélation n’est pas persistante à travers les différents échantillons, ce trait - qui est à l’origine une variable alphabétique de position - engendre d’autres interrogations, notamment celle de sa fusion par reconsidération de sa gradation. Nous verrons ceci lors de la prochaine étape. Il s’agit avant tout, ici, d’une étape de vérification statistique de l’expression bilatérale des traits non métriques dentaires que nous intégrons dans notre travail. Pour ces différentes raisons, nous choisissons donc de conserver l’hypothèse 0 pour toutes les variables avec le risque de commettre une erreur de type II (conclure de manière incorrecte que les expressions antimériques sont interdépendantes). 135 6.2.2. Le traitement des antimères Nous avons vu qu’il existe différents moyens pour traiter l’expression bilatérale. Une méthode consiste à considérer les expressions sur les dents controlatérales indépendantes l’une de l’autre et, donc, à conserver séparément les données bilatérales (tooth count method). Ce procédé ne se justifie pas, puisque la mesure de dépendance des antimères a démontré que les expressions sont fortement liées et, par conséquent, cela produit une redondance d’informations lors des analyses. Une deuxième méthode vise à retenir ou à observer uniquement l’expression d’un antimère - gauche ou droite - (lateral count method). Pour C.R. Scott et C.G. Turner (1997), ce moyen réduit la taille de l’échantillon et c’est ce que l’on cherche, en général, à éviter dans les analyses. Une troisième méthode concerne le comptage individuel (individual count), celui-ci permet de réduire les expressions antimériques par une seule valeur. Il peut être réalisé de trois façons différentes. La première réalise une moyenne des observations bilatérales, la deuxième conserve l’expression la plus basse et la dernière maintient la plus élevée. Ces trois procédés n’engendrent pas de biais majeurs dans les analyses en considérant les taux élevés de symétrie des observations controlatérales. Pour C.G. Scott et C.G. Turner, néanmoins, the antimere that shows a lower degree of expression or even trait absence is thought to reflect the action of environmental factors that do not allow the tooth to express its full genetic potential for that trait (Scott et Turner 1997, p. 103). Le traitement des observations bilatérales est une phase complexe car, selon le type de variables numériques ou alphabétisées - ou selon la composition des échantillons - dents isolées ou sujets individualisés -, il ne sera pas réalisé de la même manière. Le type de variables Des choix concernant la gradation de certaines variables sont nécessaires afin de pouvoir effectuer le traitement antimérique. Ainsi, les caractères gradués mais non ordonnés ou d’origine alphabétique, soit les variables dont leur gradation traduit une forme ou une position, font l’objet de ce type de manipulations. Pour les dents permanentes, les traits concernés sont : • Le degré de rotation (T9) Cette variable, qui consiste à juger de la rotation des incisives centrales supérieures, ne s’enregistre que si les antimères sont tous deux présents. Ils possèdent donc le même stade et leur fusion ne génère aucun conflit. Le comptage individuel est la procédure choisie pour ce trait. • Le sillon traversant (T11) Ce trait, dont il faut définir la présence et la position d’un sillon sur la face linguale des incisives supérieures, fait ici l’objet d’une dichotomisation (0-1). Ainsi, nous retenons uniquement la présence du sillon sans spécification de sa position (mésiale, distale, médiane ou mésio-distale). Cette variable peut alors faire l’objet d’une fusion par comptage individuel. • Le dessin du sillon (T53) Ce caractère décrit la forme du sillon de la surface occlusale des molaires inférieures selon les zones de contact entre les cuspides. Certains auteurs, comme K. Alt (1997) ou S. Eades (2003), remplacent l’expression antimérique asymétrique par la plus rare des expressions dans la population. Nous choisissons l’option du décompte unilatéral pour le traitement antimérique de cette variable. Nous profitons également pour repenser la gradation problématique de l’entoconulide ou 6e cuspide : • L’entoconulide (T56) Les données, pour ce trait, vont être fusionnées avec celles de l’hypoconulide (T55). L’enregistrement de l’hypoconulide consiste à définir la taille de la cinquième cuspide. L’entoconulide, s’observe uniquement en présence de six cuspides. Il s’agit d’une estimation de la taille respective des cuspides 5 et 6. Lors de la présence de la sixième cuspide, l’hypoconulide n’est pas enregistré pour éviter d’introduire une 136 Le traitement préliminaire des données information redondante. Ainsi, la fusion de ces deux traits n’engendre aucun biais méthodologique et donne naissance à la variable T55-56 dont les nouvelles gradations sont les suivantes : 0. Absence de la 5e cuspide. 1. Cuspide 5 très petite. 2. Cuspide 5 petite. 3. Cuspide 5 moyenne. 4. Cuspide 5 bien développée. 5. Cuspide 5 de taille égale aux cuspides 1 à 4. 6. Six cuspides sont présentes, la cuspide 6 est plus petite que la 5. 7. Six cuspides sont présentes, les cuspides 5 et 6 sont de taille égale. 8. Six cuspides sont présentes, la cuspide 6 est plus grande que la 5. Le comptage individuel est le procédé choisi pour cette variable. Pour les dents déciduales, les traits concernés sont : • La dilacération (T3) Cette variable décrit la courbure de la racine des incisives. Nous retenons, pour le traitement antimérique de cette variable, la méthode du décompte unilatéral. • La forme des incisives (T11 et T36) Ce trait concerne la forme de la surface incisale de l’ensemble des incisives. Nous adoptons également, pour le traitement antimérique de cette variable, la procédure du décompte unilatéral. Seuls ces six caractères font l’objet d’une modification de leur gradation ou alors d’une décision générale de leur traitement antimérique (indépendant de la composition des populations étudiées pour les variables qui font ici l’objet du comptage latéral). La composition des échantillons Le traitement antimérique diffère également selon la composition des ensembles. Les régions étudiées présentent des situations des plus diverses et souvent mixtes. Lorsque les échantillons présentent, en majorité, des dents en place (ou des sujets individualisés), les caractères sont fusionnés selon la méthode du comptage individuel (individual count) mise en place par C.G. Turner et C.R. Scott (1977) (figure 57). Nous retenons ici l’expression la plus forte lorsque les observations sont bilatérales. Les dents isolées des ensembles qui font l’objet de ce type de traitement sont alors éliminées. Type d’observation L’observation bilatérale à expression symétrique L’observation bilatérale à expression asymétrique L’observation unilatérale Méthodologie suivie Une même expression est observée, ce sera celle conservée Deux expressions différentes sont présentes, la plus forte sera retenue Une expression unique est observée et sera maintenue Figure 57 : La procédure suivie pour la fusion des données antimériques lors du comptage individuel individual count - mis en place par C.G. Turner et C.R. Scott (1977). Lorsque les échantillons sont constitués principalement de dents isolées, les variables antimériques ne sont pas fusionnées. Nous choisissons de procéder au comptage latéral (sélection aléatoire des observations gauches), sans fusion des expressions bilatérales présentes 33. Nous optons pour cette solution, car il s’agirait de travailler manuellement, entrée après entrée et variable après variable, sur des milliers d’enregistrements. Cette gestion fastidieuse des données amènerait probablement peu d’informations pertinentes supplémentaires. Nous allons maintenant définir brièvement la procédure adoptée pour chaque région, voire pour chaque site. 33 On se réfère aux observations bilatérales asymétriques qui peuvent fournir des indications complémentaires. Les données antimériques symétriques, quant à elles, ne fournissent dans ce cas précis aucune information supplémentaire. 137 Chapitre 6 6.2.2.1. Le traitement des antimères pour la Bohême La Bohême est la région présentant le nombre de sites le plus élevé (n=64). Les ensembles sont constitués en majorité de sépultures individuelles, facilitant ainsi le traitement des données bilatérales puisque chaque sujet est identifié et les échantillons ne comportent pas de dents isolées. Le traitement des antimères a été réalisé sur les dents permanentes et déciduales. Nous avons effectué la fusion des expressions antimériques par la méthode du comptage individuel pour chaque variable, à l’exception des quelques traits qui font l’objet du comptage latéral (T53 pour la dentition permanente, T3, T11 et T36 pour la dentition déciduale). 6.2.2.2. Le traitement des antimères pour la Hongrie La Hongrie est représentée par quinze sites. Les sépultures, composées d’inhumations et d’incinérations, sont individuelles. Le traitement des données bilatérales s’effectue sur des sujets identifiés sans dents isolées. La procédure s’effectue uniquement sur la dentition permanente, celle déciduale ayant été éliminée. Nous avons réalisé la fusion des expressions antimériques par la méthode du comptage individuel pour chaque variable, à l’exception de la variable T53 qui fait l’objet du comptage latéral. 6.2.2.3. Le traitement des antimères pour le sud de la France Le sud de la France comporte cinq sépultures collectives dont certaines avec plusieurs phases d’occupations. Ces ensembles sont, en grande majorité, composés de dents isolées, à l’exception de l’hypogée des Boileau pour laquelle les individus sont identifiés. Cette situation mixte nous contraint à travailler selon la composition des sites. Le traitement des antimères a été réalisé sur les dents permanentes et déciduales. Ainsi, pour l’hypogée des Boileau (F-3), nous avons procédé à la fusion des expressions antimériques par la méthode du comptage individuel pour chaque variable, à l’exception des quelques traits qui font l’objet du comptage latéral (T53 pour la dentition permanente, T3, T11 et T36 pour la dentition déciduale). Les autres sépultures - le dolmen des Peirières (F-1), le dolmen des Peyraoutes (F-2), l’hypogée des Crottes (F-4) et le dolmen des Cudières (F-5) -, ont été traitées selon la méthode du comptage latéral. 6.2.2.4. Le traitement des antimères pour le nord de l’Espagne Le nord de l’Espagne, constitué par quatorze sites, présente une situation composite avec des sépultures individuelles et collectives. Nous optons pour un traitement selon la composition majeure de chaque site. La procédure s’effectue uniquement sur la dentition permanente. Nous avons réalisé, pour les ensembles ci-dessous, la fusion des expressions antimériques par la méthode du comptage individuel pour chaque variable, à l’exception de la variable T53 qui fait l’objet du comptage latéral. Les dents non attribuées ont été éliminées. • Carrelasvegas (ES-1) • La Sima (ES-8) • Cerro de la Cabeza (ES-2) • Los Cercados (ES-11) • Cueva Rubia, (ES-4) • Los Zumacalles (ES-12) • Cueva Tino (ES-5) • • El Tomillar (ES-6) Les différentes grottes de la Valle de las Higueras (ES-13) • Villimar (ES-14) • Fuente Olmedo (ES-7) Nous avons traité, les ensembles ci-dessous, selon la procédure du comptage latéral. 138 Le traitement préliminaire des données • La Cueva del Destete (ES-3) • La Vellila (ES-9) • Las Arnillas (ES-10) • le tumulus de la Valle de Las Higueras (ES-13) 6.2.2.5. Le traitement des antimères pour la Suisse La Suisse révèle, selon la période culturelle envisagée, une situation mixte. Ainsi, comme pour les deux régions précédentes, nous l’avons abordée selon la composition majoritaire des ensembles. Le traitement des antimères a été réalisé sur les dents permanentes et déciduales. Nous avons procédé, pour les échantillons listés ci-dessous, à la fusion des expressions antimériques par la méthode du comptage individuel pour chaque variable, à l’exception des quelques traits qui font l’objet du comptage latéral (T53 pour la dentition permanente, T3, T11 et T36 pour la dentition déciduale). Les dents non attribuées ont été éliminées. • Barmaz I (CH-1) • Corseaux (CH-6) • Barmaz II (CH-2) • • Chamblandes (CH-3) • Petit-Chasseur (CH-7) : l’occupation campaniforme du dolmen MVI et le dolmen MXI Chemin des Collines (CH-5) • Petit-Chasseur (CH-7) : le dolmen MXII, l’occupation néolithique récent du dolmen MVI et la fosse à incinération du dolmen MVI • Chlus-Tschäpperli (CH-4) • Avenue Ritz (CH-8) Nous avons traité les ensembles ci-dessous, composés pour une large majorité de dents isolées, selon la procédure du comptage latéral. 6.2.2.6. Le traitement des antimères : un bilan Le traitement des données bilatérales vise principalement à éviter d’introduire des informations redondantes au sein des analyses. En effet, la mesure de la dépendance entre les antimères s’est révélée positive pour une très large majorité de traits que nous avons observés. Deux méthodes ont été retenues : • Le comptage individuel: fusion des données bilatérales selon la méthode mise en place par C.G. Turner et C.R. Scott (1977). • Le comptage latéral : choix arbitraire de l’expression d’un antimère uniquement (les données gauches ici). Nous n’avons pas choisi un traitement par région étudiée. La Bohême et la Hongrie, composées toutes deux de sujets identifiés, ont fait l’objet d’un traitement par comptage individuel. Les échantillons provenant de régions présentant des situations composites, - le sud de la France, le nord de l’Espagne et la Suisse -, ont été traités de cas en cas. Les sites, majoritairement composés de dents isolées, ont fait l’objet d’un comptage latéral. La procédure du comptage individuel a été réalisée pour les ensembles constitués principalement de sépultures permettant une individualisation des sujets. Les dents isolées de ces échantillons ont alors été éliminées afin de ne pas augmenter faussement les corpus. La fusion des antimères permet d’améliorer les effectifs des populations. De ce fait, nous avons choisi de travailler de cette manière pour les régions mixtes afin de ne pas pénaliser les sites qui pouvaient faire l’objet d’une optimisation par comptage individuel. 139 Chapitre 6 6.3. Les corrélations entre les traits Cette étape du traitement préliminaire consiste en l’évaluation des relations entre les différentes variables. L’hypothèse de départ est de considérer les expressions des caractères dentaires indépendantes les unes des autres, néanmoins on ne se réfère pas à un trait exprimé à l’intérieur d’une même classe morphologique. C’est pourquoi, nous allons aborder ici brièvement ces deux éléments. L’effet du champ morphogénétique Nous avons vu, dans le chapitre concernant les aspects biologiques de la dentition (c.f. chapitre 4.2.1.3.), que cette dernière présente des gradients où chaque dent ressemble à sa voisine. Ce continuum morphologique, allant de l’incisive à la molaire, se retrouve également au sein des différentes classes dentaires. Et c’est ce dernier point qui nous importe ici. Le terme de gradient d’expression est employé pour comprendre le comportement d’une variable au sein d’une classe dentaire. Lorsqu’un trait est présent sur un membre d’un district, il est fort probable de l’observer sur l’ensemble du groupe. Les chercheurs expliquent ce phénomène en terme de champs morphogénétiques (Butler 1939) et de clones (Osborn 1978). L’intensité de l’influence morphogénétique est meilleure sur une dent particulière dans chaque classe, et cette dent est la plus stable dans sa taille et sa forme. Au sein d’un district, les dents en position mésiale posséderaient une plus grande concentration en substance et seraient celles soumises au plus fort contrôle génétique. Au contraire, les dents en position distale seraient celles qui montreraient les plus grandes composantes mésologiques. Prenons, par exemple, l’expression de l’hypocone sur les molaires supérieures (T26 - quatrième cuspide), nous avons de grandes chances d’observer ce trait sur les trois dents. La taille de l’hypocone de la deuxième molaire, d’après le modèle attendu du gradient d’expression, devrait être intermédiaire entre celle de la première et celle de la troisième. L’interdépendance d’un trait au sein d’une classe dentaire ne fait que peu de doutes, même si elle n’est pas toujours parfaite. Certains chercheurs, comme C.R. Scott et C.G. Turner (1997), considèrent que les données exploitables d’une classe dentaire sont différentes selon le type d’étude envisagé. Lors d’une caractérisation de populations, il est possible de considérer l’ensemble des fréquences des différents membres d’un district dentaire. En revanche, lors d’études impliquant des distances entre populations, il est souhaitable de ne retenir qu’un seul membre par classe dentaire, qu’ils nomment dent clé (figure 58). District Dent clé - maxillaire Dent clé - mandibule Incisive permanente Canine permanente I1 C P1 pour les traits sur les racines P2 pour les traits sur les couronnes M1 i1 c m1, m2 I2 C P1 pour les traits sur les couronnes P2 pour les traits sur les racines M1 i2 c m1, m2 Prémolaire permanente Molaire permanente Incisive déciduale Canine déciduale Molaire déciduale Figure 58 : Les dents clés fournissant selon C.R. Scott et C.G. Turner (1997) les meilleures informations dans les études entre populations. Il n’y a pas de règles préétablies concernant les données qui peuvent être mobilisées au sein d’un district. Certaines études mesurent le degré de dépendance des variables intradistrict (Eades 1997, 2003, Desideri 2001, à paraître...), d’autres ne semblent pas le faire (Irish 1998, Coppa et al. 1998). Certains auteurs emploient l’ensemble des variables (Coppa et al. 1998, Parras 2004), d’autres fusionnent les données provenant des différents membres des classes dentaires (Eades 2003), ou encore ne retiennent que la dent clé (Turner 1967, Irish 1998, Desideri 2001, à paraître...). 140 Le traitement préliminaire des données L’indépendance des variables L’inter-indépendance est envisagée au niveau des différentes variables (Scott 1975). Les recherches reposent essentiellement sur les travaux de C.R. Scott (1975, 1978, 1979) qui a testé un grand nombre de traits non métriques dentaires. Des études rapportent ponctuellement certaines associations entre variables34, qui ne se révèlent pas persistantes à travers les différentes populations. Selon C.G. Turner, one (correlation) that persists at a low positive level is Carabelli and protostylid, as well as the long known correlation between congenitally missing teeth and third molar size reduction or absence. Our many years of global comparisons lead us to believe that most of our key traits are truly independant variables, or as much so as features can ever be developing within a single organ (C.G. Turner, communication personnelle). Cette partie du traitement préliminaire des données sert principalement à estimer certains types de relations entre les variables dans une perspective de sélection des variables que nous allons intégrer dans les analyses. Elle s’effectue en trois temps : • Les corrélations intradistrict Il s’agit ici de vérifier si l’expression intradistrict des variables dentaires suit ou non le modèle du gradient d’expression en vue des données à intégrer lors des analyses. Notre intuition, après avoir observé des milliers de dents, est qu’une très large majorité de variables réagissent selon ce modèle. Selon les populations, les mesures reposeront sur des effectifs limités, c’est pourquoi une prudence particulière doit être envisagée lors de l’interprétation des résultats. • Les corrélations interdistrict Il s’agit d’une extension du point précédent. Certains traits s’expriment au delà d’une classe dentaire et nous souhaitons évaluer les relations d’un trait entre les membres des différents districts. Selon les décisions prises pour les corrélations intradistrict et les résultats de cette mesure, avec les réserves nécessaires selon les régions étudiées, il sera peut-être envisagé de devoir sélectionner une dent clé sur l’ensemble des classes dentaires. • La corrélation intertrait : trait de Carabelli (T24) - protostylide (T54) Nous mesurons ici uniquement la dépendance ou non de deux variables exprimées sur les molaires permanentes, soit le trait de Carabelli (T24) s’exprimant sur les molaires supérieures et le protostylide (T54) s’observant sur les inférieures puisque, selon C.G. Turner (c.f. ci-dessus), cette association serait la plus souvent rencontrée dans les différentes études indépendamment des populations analysées. Nous avons délibérément choisi de ne pas traiter l’ensemble des variables pour deux raisons très simples. Premièrement, plusieurs travaux (Eades 1997, Desideri 2001…) ont illustré la difficulté du maniement et de l’interprétation de matrices de corrélations incluant un grand nombre de variables. Deuxièmement, les corrélations sont souvent importantes sans pour autant avoir une réelle signification biologique (C.G. Turner, communication personnelle). La difficulté ici sera de réaliser ces mesures sur des échantillons constitués pour la plupart de dents isolées et, par conséquent, fournissant des informations que nous devrons considérer avec prudence. Ainsi, deux régions auront un plus grand poids dans les décisions concernant la mobilisation des données pour les analyses : la Bohême et la Suisse. 34 Les associations entre variables les plus citées dans la littérature sont : le tubercule de Carabelli et le protostylide (Scott 1978) - le tubercule de Carabelli et l’hypocone (Keene 1968, Scott 1979) - les crêtes linguales marginales et la crête accessoire distale de la canine (Dahlberg 1963, Scott 1975) - l’odontome et les cuspides accessoires des prémolaires (Turner et al. 1991) - le tubercule dentaire des incisives et des canines (Berry 1963). 141 Chapitre 6 6.3.1. Les corrélations intradistrict Nous souhaitons mettre en évidence les relations existant entre les expressions d’une variable sur les dents appartenant au même district. Nous l’effectuons par une mesure non paramétrique d’association pour variables ordinales ou graduées, le Tau de Kendall. Nous déterminons les hypothèses de départ suivantes : H0 = hypothèse nulle : les expressions d’une variable observée sur les dents d’une même classe dentaire sont corrélées, elles suivent le modèle attendu : l’effet du champ morphogénétique. H1 = hypothèse alternative : les expressions d’une variable observée sur les dents d’une même classe dentaire ne sont pas corrélées, elles ne suivent pas le modèle attendu. Les hypothèses sont testées au seuil de signification de 0.05 ou 0.01. Selon le seuil de signification retenu, les expressions sont significativement corrélées si leur valeur est inférieure au seuil. Le calcul des corrélations s’effectue sur 86 groupes de variables, dont 55 sur la dentition permanente et 31 sur celle déciduale (figure 59). DENTS PERMANENTES Maxillaire M3 M2 M1 P2 P1 C I2 I1 1 Absence congénitale 2 Nombre de radicules 3 Racine appendiciforme 4 Nombre de racines des prémolaires 5 Nombre de racines des molaires 6 Racine de Carabelli 7 Racine paramolaire 8 Racine idiopathique 9 Degré de rotation 10 Convexité vestibulaire 11 Sillon traversant 12 Tubercule distal 13 Cingulum vestibulaire 14 Gouttière vestibulaire 15 Crêtes linguales marginales 16 Tubercule dentaire 17 Crêtes vestibulaires marginales 18 Crête mésiale 19 Crête accessoire distale 20 Cuspides accessoires 21 Odontome 22 Extension de l'émail 23 Parastyle 24 Trait de Carabelli 25 Métacone 26 Hypocone 27 Métaconule 28 Tubercule distal accessoire 29 Tubercule mésial paracone 30 Tubercule mésial accessoire 31 Protoconule 32 Tubercule lingual paracone Mandibule 33 Absence congénitale 34 Nombre de radicules 35 Racine appendiciforme 36 Nombre racines canines 37 Racine de Tomes 38 Nombre racines molaires 39 Racine de Citroen 40 Racine paramolaire 41 Racine entomolaire 42 Racine idiopathique 43 Crêtes linguales marginales 44 Crêtes vestibulaires marginales 45 Crête accessoire distale 46 Odontome 47 Variation des cuspides linguales 48 Pli cuspide mésiolinguale 49 Fosse précuspidale 50 Crête trigonide intermédiaire 51 Crête trigonide distale 52 Nombre de cuspides 53 Dessin du sillon 54 Protostylide 55-56 Hypoconulide-Entoconulide 57 Métaconulide 58 Tubercule paracone 59 Tubercule de Citroen 142 DENTS DECIDUALES Maxillaire m2 m1 c i2 i1 m2 m1 c i2 i1 1 Nombre de radicules 2 Extension de l'émail 3 Dilacération 4 Racine appendiciforme 5 Nombre de racines 6 Racine de la canine 7 Nombre de racines molaires 8 Racine de Carabelli 9 Racine paramolaire 10 Racine idiopathique 11 Forme des incisives 12 Gouttière vestibulaire 13 Crêtes linguales marginales 14 Tubercule dentaire 15 Développement cuspides 16 Crête du métacone 17 Parastyle 18 Trait de Carabelli 19 Métacone 20 Hypocone 21 Métaconule 22 Tubercule distal accessoire 23 Tubercule mésial paracone 24 Tubercule mésial accessoire 25 Protoconule 26 Tubercule lingual paracone M3 M2 M1 P2 P1 C I2 I1 Mandibule 27 Nombre de radicules 28 Extension de l'émail 29 Racine appendiciforme 30 Nombre de racines 31 Nombre de racines molaires 32 Racine de Citroen 33 Racine paramolaire 34 Racine idiopathique 35 Racine entomolaire 36 Forme des incisives 37 Double tubercule lingual 38 Développement cuspides 39 Protostylide 40 Fosse précuspidale 41 Distostylide fissuré 42 Crête trigonide intermédiaire 43 Crête trigonide distale 44 Pli mésiolingual 45 Hypoconulide 46 Entoconulide 47 Métaconulide 48 Tubercule paracone 49 Tubercule de Citroen 50 Fosse postérieure 51 Sillon trigonide 52 Proéminence cervicale Figure 59 : Représentation synthétique concernant les groupes de variables testés pour la mesure de dépendance intradistrict. En gris foncé, les variables éliminées lors des phases antérieures du traitement préliminaire des données. Le traitement préliminaire des données 6.3.1.1. Les corrélations intradistrict pour la Bohême Cette mesure a pu être réalisée pour la Bohême sur 80% (n=44) des groupes testés pour la dentition permanente et 32% (n=10) pour la dentition déciduale. Les résultats se trouvent en table 16. Les calculs de corrélations montrent que les variables s’exprimant sur la dentition permanente suivent à plus de 93% (n=41) l’effet du champ. Seuls trois traits ne présentent pas de corrélations intradistrict significatives. Il s’agit du sillon traversant (T11), du métaconulide (T57) et du tubercule paracone (T58). La situation n’est pas la même pour la dentition déciduale. En effet, les groupes de variables qui ont fait l’objet d’un calcul (n=10) n’ont dévoilé aucune corrélation significative, mais les effectifs des échantillons de la dentition temporaire pourraient être à l’origine de ces résultats plutôt surprenants. 6.3.1.2. Les corrélations intradistrict pour la Hongrie Cette mesure a pu être réalisée pour la Hongrie sur 33% (n=18) des groupes testés pour la dentition permanente. Les résultats se trouvent en table 17. Aucun calcul n’est effectué sur la dentition déciduale puisqu’elle a été éliminée lors d’une phase précédant le traitement préliminaire. Les résultats des corrélations montrent que plus de 61% des traits s’exprimant sur la dentition définitive suivent l’effet du champ. Sept variables ne présentent pas de corrélations intradistrict significatives. Il s’agit du nombre de radicules des molaires (T2), du nombre de racines des prémolaires (T4), des crêtes linguales marginales (T15), du métacone (T25), de l’hypocone (T26), du tubercule accessoire distal (T28) et du dessin du sillon (T53). 6.3.1.3. Les corrélations intradistrict pour le sud de la France Cette mesure a pu être réalisée pour cette région sur 45% (n=25) des groupes testés pour la dentition permanente et 10% seulement (n=3) pour la dentition déciduale. Les résultats se trouvent en table 18. Les calculs des corrélations montrent que les variables s’exprimant sur la dentition permanente suivent à plus de 68% (n=25) l’effet du champ. Huit traits ne présentent pas de corrélations intradistrict significatives. Il s’agit du nombre de racines des prémolaires (T4), du nombre de racines des molaires (T5), des crêtes linguales marginales (T15), du tubercule dentaire (T16), du tubercule accessoire distal (T28), du métaconule (T27), du tubercule mésial paracone (T29) et du métaconulide (T57). La situation pour la dentition déciduale est comparable aux résultats pour la Bohême. En effet, peu de variables ont fait l’objet d’un calcul (n=3) et aucune d’entre elles n’a présenté une corrélation significative. Les effectifs des échantillons sont ici sans aucun doute à l’origine de ces résultats médiocres. 6.3.1.4. Les corrélations intradistrict pour le nord de l’Espagne Cette mesure a pu être réalisée pour le nord de l’Espagne sur 33% (n=18) des groupes testés pour la dentition permanente. Les résultats se trouvent en table 19. Aucun calcul n’est effectué sur la dentition déciduale puisqu’elle a été éliminée lors d’une phase précédente du traitement préliminaire. Les calculs des corrélations montrent que les variables s’exprimant sur la dentition permanente suivent à 44% (n=8) seulement l’effet du champ. Dix traits ne présentent pas de corrélations intradistrict significatives. Il s’agit du nombre de racines des prémolaires (T4), du nombre de racines des molaires (T5), de la convexité vestibulaire (T10), des crêtes linguales marginales (T15), de l’extension de l’émail des molaires (T22), du tubercule accessoire distal (T28), du nombre de cuspides (T52), du dessin du sillon (T53) et de l’hypoconulide-entoconulide (T55-56). 143 Chapitre 6 6.3.1.5. Les corrélations intradistrict pour la Suisse Cette mesure a pu être réalisée pour la Suisse sur 77% (n=37) des groupes testés pour la dentition permanente et 46% (n=14) pour la dentition déciduale. Les résultats se trouvent en table 20. Les calculs des corrélations montrent que les variables s’exprimant sur la dentition permanente suivent à plus de 76% (n=37) l’effet du champ. Neuf traits ne présentent pas de corrélations intradistrict significatives. Il s’agit du nombre de radicules des incisives (T2), des crêtes linguales marginales (T15), des cuspides accessoires (T20), du parastyle (T23), du tubercule mésial paracone (T29), du tubercule lingual paracone (T32), des variations des cuspides linguales (T47), du dessin du sillon (T53) et du métaconulide (T57). Pour la dentition déciduale, les variables présentant des corrélations significatives s’élèvent à plus de 79% (n=11). Trois traits ne présentent pas de corrélations intradistrict significatives. Il s’agit de la forme des incisives supérieures (T11) et inférieures (T36) ainsi que du développement des cuspides (T38). Nous avons des résultats basés sur des effectifs raisonnables uniquement pour cette région et d’une manière générale les variables semblent suivre le modèle attendu. 6.3.1.6. Les corrélations intradistrict : un bilan Cette partie du traitement de la dépendance de l’expression des variables au sein d’un même district est intéressante puisque les résultats ne sont pas aussi évidents et les réponses varient selon les échantillons analysés. Nous décidons que : • si un trait se révèle non corrélé sur les différents membres d’un district pour les deux régions majeures (Bohême et Suisse) ou alors pour une région majeure et au moins deux des régions mineures, nous retenons l’hypothèse 1 pour cette variable avec le risque d’erreur de commettre une erreur de type I (conclure de manière erronée que les expressions d’une variable au sein d’une même classe dentaire ne suivent pas le gradient d’expression). • pour toutes les autres configurations, nous conservons l’hypothèse 0 avec le risque de commettre une erreur de type II (conclure de manière incorrecte que les expressions d’une variable au sein d’une même classe dentaire suivent le gradient d’expression). Seules trois variables, selon nos critères, répondent à l’exigence H1: - les crêtes linguales marginales (T15) - le dessin du sillon (T53) - le métaconulide (T57) Ainsi, pour ces traits, l’ensemble des membres peuvent être sélectionnées pour les analyses. En revanche, pour toutes les autres variables, seule la dent clé, définie par C.R. Scott et C.G. Turner (1997), devrait être retenue. 144 Le traitement préliminaire des données 6.3.2. Les corrélations interdistrict Nous souhaitons mettre en évidence les relations entre les expressions d’une variable sur les dents appartenant à des districts différents. Nous l’effectuons par une mesure non paramétrique d’association pour variables ordinales ou graduées, le Tau de Kendall. Nous déterminons les hypothèses de départ suivantes : H0 = hypothèse nulle : les expressions d’une variable observée sur les dents de classes dentaires différentes sont corrélées. H1 = hypothèse alternative : les expressions d’une variable observée sur les dents de classes dentaires différentes ne sont pas corrélées. Les hypothèses sont testées au seuil de signification de 0.05 ou 0.01. Selon le seuil de signification retenu, les expressions sont significativement corrélées si leur valeur est inférieure au seuil. Le calcul des corrélations s’effectue sur 11 groupes de variables, dont 7 sur la dentition permanente et 4 sur celle déciduale (figure 60). Cette mesure ne concerne pas les variables exprimées sur les racines. Nous avons choisi de ne pas les tester puisque ces traits ne présentent que très peu de variations. DENTS PERMANENTES Maxillaire M3 M2 M1 P2 P1 C I2 I1 1 Absence congénitale 2 Nombre de radicules 3 Racine appendiciforme 4 Nombre de racines des prémolaires 5 Nombre de racines des molaires 6 Racine de Carabelli 7 Racine paramolaire 8 Racine idiopathique 9 Degré de rotation 10 Convexité vestibulaire 11 Sillon traversant 12 Tubercule distal 13 Cingulum vestibulaire 14 Gouttière vestibulaire 15 Crêtes linguales marginales 16 Tubercule dentaire 17 Crêtes vestibulaires marginales 18 Crête mésiale 19 Crête accessoire distale 20 Cuspides accessoires 21 Odontome 22 Extension de l'émail 23 Parastyle 24 Trait de Carabelli 25 Métacone 26 Hypocone 27 Métaconule 28 Tubercule distal accessoire 29 Tubercule mésial paracone 30 Tubercule mésial accessoire 31 Protoconule 32 Tubercule lingual paracone Mandibule 33 Absence congénitale 34 Nombre de radicules 35 Racine appendiciforme 36 Nombre racines canines 37 Racine de Tomes 38 Nombre racines molaires 39 Racine de Citroen 40 Racine paramolaire 41 Racine entomolaire 42 Racine idiopathique 43 Crêtes linguales marginales 44 Crêtes vestibulaires marginales 45 Crête accessoire distale 46 Odontome 47 Variation des cuspides linguales 48 Pli cuspide mésiolinguale 49 Fosse précuspidale 50 Crête trigonide intermédiaire 51 Crête trigonide distale 52 Nombre de cuspides 53 Dessin du sillon 54 Protostylide 55 Hypoconulide 55-56 Hypoconulide-Entoconulide 58 Tubercule paracone 59 Tubercule de Citroen DENTS DECIDUALES Maxillaire m2 m1 c i2 i1 m2 m1 c i2 i1 1 Nombre de radicules 2 Extension de l'émail 3 Dilacération 4 Racine appendiciforme 5 Nombre de racines 6 Racine de la canine 7 Nombre de racines molaires 8 Racine de Carabelli 9 Racine paramolaire 10 Racine idiopathique 11 Forme des incisives 12 Gouttière vestibulaire 13 Crêtes linguales marginales 14 Tubercule dentaire 15 Développement cuspides 16 Crête du métacone 17 Parastyle 18 Trait de Carabelli 19 Métacone 20 Hypocone 21 Métaconule 22 Tubercule distal accessoire 23 Tubercule mésial paracone 24 Tubercule mésial accessoire 25 Protoconule 26 Tubercule lingual paracone M3 M2 M1 P2 P1 C I2 I1 Mandibule 27 Nombre de radicules 28 Extension de l'émail 29 Racine appendiciforme 30 Nombre de racines 31 Nombre de racines molaires 32 Racine de Citroen 33 Racine paramolaire 34 Racine idiopathique 35 Racine entomolaire 36 Forme des incisives 37 Double tubercule lingual 38 Développement cuspides 39 Protostylide 40 Fosse précuspidale 41 Distostylide fissuré 42 Crête trigonide intermédiaire 43 Crête trigonide distale 44 Pli mésiolingual 45 Hypoconulide 46 Entoconulide 47 Métaconulide 48 Tubercule paracone 49 Tubercule de Citroen 50 Fosse postérieure 51 Sillon trigonide 52 Proéminence cervicale Figure 60 : Représentation synthétique concernant les groupes de variables testés pour la mesure de dépendance interdistrict. En gris foncé, les variables éliminées lors des phases antérieures du traitement préliminaire des données. 145 Chapitre 6 6.3.2.1. Les corrélations interdistrict pour la Bohême Pour la Bohême, cette mesure a été réalisée sur six des huit groupes de la dentition permanente et sur un seul des quatre groupes de la dentition déciduale. Les résultats se trouvent en table 21. En ce qui concerne les dents permanentes, seul le tubercule dentaire (T16) des incisives et des canines montre une corrélation significative entre les différents membres des deux districts. Les crêtes linguales marginales (T15) se révèlent partiellement significatives, soit uniquement la corrélation entre l’incisive centrale et la canine. Les associations restantes ne se sont pas révélées significatives. Pour la dentition déciduale, une seule mesure (T13 - les crêtes linguales marginales -) non corrélée a été effectuée. 6.3.2.2. Les corrélations interdistrict pour la Hongrie Pour la Hongrie, cette mesure a été réalisée sur cinq des huit groupes de la dentition permanente. Les résultats se trouvent en table 22. Aucun calcul n’est effectué sur la dentition déciduale puisqu’elle a été éliminée lors d’une phase précédente du traitement préliminaire. Trois variables présentent des corrélations significatives partielles. Il s’agit de l’absence congénitale (T1) des troisièmes molaires et des incisives latérales, du tubercule dentaire (T16) des incisives et des canines et des crêtes vestibulaires marginales (T17) des canines et des prémolaires. 6.3.2.3. Les corrélations interdistrict pour le sud de la France Pour le sud de la France, cette mesure a été réalisée sur trois des huit groupes de la dentition permanente et aucun de ceux définis pour la dentition déciduale n’ont été calculés. Les résultats se trouvent en table 23. Une seule des associations calculées s’est révélée partiellement significative, soit le tubercule dentaire (T16) des incisives latérales et des canines. 6.3.2.4. Les corrélations interdistrict pour le nord de l’Espagne Pour le nord de l’Espagne, cette mesure a été réalisée sur trois des huit groupes de la dentition permanente seulement. Les résultats se trouvent en table 24. Aucun calcul n’est effectué sur la dentition déciduale puisqu’elle a été éliminée lors d’une phase précédente du traitement préliminaire. Les crêtes linguales marginales (T15) des incisives centrales et des canines et les crêtes vestibulaires marginales (T17) des canines et des prémolaires présentent des corrélations significatives partielles. 6.3.2.5. Les corrélations interdistrict pour la Suisse Pour la Suisse, cette mesure a été réalisée sur sept des huit groupes de la dentition permanente et sur deux des quatre groupes de la dentition déciduale. Les résultats se trouvent en table 25. Trois variables présentent des corrélations significatives, dont deux partielles. Il s’agit de l’absence congénitale (T1) des molaires et des incisives latérales, du tubercule dentaire (T16) des incisives et des canines et des crêtes vestibulaires marginales (T17) des incisives latérales et des prémolaires. Pour la dentition déciduale, seules les crêtes linguales marginales (T13) se sont révélées corrélées. 6.3.2.6. Les corrélations interdistrict : un bilan Il s’est agi de tester l’interdépendance d’une variable s’exprimant au delà d’un district dentaire. L’objectif étant de définir si l’effet du champ peut également être mis en évidence sur plusieurs classes dentaires. Les réponses sont à nouveau variables selon les échantillons analysés. La procédure définie pour les corrélations intradistrict va être à nouveau employée. 146 Le traitement préliminaire des données Nous décidons que : • si les expressions d’un trait se révèlent corrélées sur les membres de divers districts pour les deux régions majeures (Bohême et Suisse) ou alors pour une région majeure et au moins deux des régions mineures, nous retenons l’hypothèse 0 pour cette variable avec le risque d’erreur de commettre une erreur de type II (conclure de manière incorrecte que les expressions d’une variable sur les dents appartenant à des districts différents sont interdépendantes) • pour toutes les autres configurations, nous conservons l’hypothèse 1 avec le risque de commettre une erreur de type I (conclure de manière erronée que les expressions d’une variable sur les dents appartenant à des districts différents sont indépendantes) Une seule variable répond à l’exigence H0, il s’agit du tubercule dentaire (T16). Cette association, qui se révèle significative sur l’ensemble des régions étudiées - à l’exception de l’Espagne -, a également été mise en évidence dans plusieurs travaux (Berry 1963, Scott 1979). De ce fait, lors de la sélection des variables mobilisables pour les analyses, une dent clé seulement doit être retenue pour ce trait qui s’exprime tant sur les incisives que sur la canine. Pour toutes les autres variables testées, les dents clés de chaque district peuvent être maintenues. 6.3.3. Les corrélations intertrait : trait de Carabelli (T24) - protostylide (T54) Nous souhaitons tester l’association entre trait de Carabelli (T24) - protostylide (T54). Nous l’effectuons à nouveau par la mesure Tau de Kendall. Nous déterminons les hypothèses de départ suivantes : H0 = hypothèse nulle : les deux variables observées sont corrélées, les expressions du tubercule de Carabelli et du protostylide sont interdépendantes. H1 = hypothèse alternative : les deux variables observées ne sont pas corrélées, les expressions du tubercule de Carabelli et du protostylide sont indépendantes. Les hypothèses sont testées au seuil de signification de 0.05 ou 0.01. Selon le seuil de signification retenu, les expressions des variables sont significativement corrélées si leur valeur est inférieure au seuil. Les mesures pour chaque région de cette association sont regroupées en table 26. 6.3.3.1. Les corrélations intertrait pour la Bohême Trois associations significatives ont été mises en évidence pour la Bohême impliquant les trois molaires supérieures avec les deux premières molaires inférieures. Il s’agit de : - l’expression du tubercule de Carabelli de la première molaire supérieure avec l’expression du protostylide de la première molaire inférieure. - l’expression du tubercule de Carabelli de la deuxième molaire supérieure avec l’expression du protostylide de la première molaire inférieure. - l’expression du tubercule de Carabelli de la troisième molaire supérieure avec l’expression du protostylide de la deuxième molaire inférieure. 6.3.3.2. Les corrélations intertrait pour la Hongrie Pour la Hongrie, aucune corrélation significative n’a été mise en évidence entre les expressions de deux membres appartenant chacun à l’une des variables testées. 147 Chapitre 6 6.3.3.3. Les corrélations intertrait pour le sud de la France Une seule association se révèle significative pour le sud de la France. Il s’agit de l’expression du tubercule de Carabelli de la troisième molaire supérieure avec l’expression du protostylide de la deuxième molaire inférieure. 6.3.3.4. Les corrélations intertrait pour le nord de l’Espagne Le nord de l’Espagne révèle à nouveau l’association significative de l’expression du tubercule de Carabelli de la troisième molaire supérieure avec l’expression du protostylide de la deuxième molaire inférieure. 6.3.3.5. Les corrélations intertrait pour la Suisse La Suisse, comme la Hongrie, ne montre aucune corrélation significative entre les expressions de deux membres appartenant chacun à l’une des variables testées. 6.3.3.6. Les corrélations intertrait : un bilan Nous avons mesuré une seule association entre variables s’exprimant sur les arcades opposées (tubercule de Carabelli - T24 - et protostylide - T54 -). Nous décidons de suivre la procédure adoptée jusqu’ici, soit : • si les expressions de deux membres appartenant chacun à l’une des variables se révèlent corrélées pour les deux régions majeures (Bohême et Suisse) ou alors pour une région majeure et au moins deux des régions mineures, nous retenons l’hypothèse 0 pour cette association avec le risque d’erreur de commettre une erreur de type II (conclure de manière incorrecte que les expressions sont interdépendantes). • pour toutes les autres configurations, nous conservons l’hypothèse 1 avec le risque de commettre une erreur de type I (conclure de manière erronée que les expressions sont indépendantes). Une seule mesure répond à l’exigence H0, il s’agit de l’association de l’expression du tubercule de Carabelli de la troisième molaire supérieure avec l’expression du protostylide de la deuxième molaire inférieure. Cette corrélation significative n’affectera pas les traits exploitables pour les analyses puisqu’elle n’implique aucune dent clé. 148 Le traitement préliminaire des données 6.4. Le dimorphisme sexuel Les chercheurs se sont énormément intéressés à l’influence du sexe des individus sur les incidences des caractères morphologiques dentaires. Il semblerait que les gènes des chromosomes sexuels soient impliqués dans le développement de la dentition, cependant les conséquences au niveau phénotypique paraissent minimes (Scott et Turner 1997). Pour disposer d’une signification biologique évidente, les différences dans l’expression d’une variable devraient, en principe, présenter une prédisposition comparable à travers les différentes populations (Hauser et De Stefano 1989). Mais, à ce jour, aucune étude n’a démontré des divergences sexuelles pan-humaines concrètes pour les traits non métriques dentaires (Eades 2003). Des divergences dentaires se révèlent liées au sexe des individus comme la taille des dents (Garn et al. 1966a/b, 1967, Mayhall et Alvesalo 1992, Harila 2004) ou encore la fréquence d’hypodontie et d’hyperdontie (Brook 1984). Pour les variations non métriques, proprement dites, les avis diffèrent selon les auteurs. Prenons l’exemple de deux variables: le tubercule de Carabelli et les crêtes linguales marginales. Ces traits ont fait l’objet d’un certain nombre de recherches, dont la détermination du dimorphisme sexuel au sein des populations les plus diverses. Les résultats sont des plus contradictoires. En effet, selon les études, ces variables sont soit sexuellement dimorphiques (pour le tubercule de Carabelli : Goose et Lee 1971, Kaul et Prakash 1981, Kieser et Preston 1981 et pour les crêtes linguales marginales : Rothhammer et al. 1968, Harris 1980), soit elles ne montrent aucune différence significative (pour le tubercule de Carabelli : Garn et al. 1966a, Scott 1980, Townsend et Martin 1992 et pour les crêtes linguales marginales : Portin et Alvesalo 1974, Mizoguchi 1985). Les réponses peu convaincantes posent quelques difficultés. En effet, les traits sexuellement dimorphiques - variant selon les populations étudiées - ne montrent aucune constance. De plus, lorsque des différences sont enregistrées, elles se résument, la plupart du temps, à une expression plus forte chez les hommes (Scott et Turner 1997). L’expression plus prononcée pour certains traits chez les hommes est mise en relation avec la taille des dents, qui est elle-même influencée par le sexe des individus. Le diamètre mésio-distal des couronnes est la mesure qui montrerait le plus de différences sexuelles (Garn et al. 1967). Cette mesure, généralement plus élevée chez l’homme, pourrait être à l’origine d’un développement plus prononcé de la morphologie (Tyrell 2000). La dépendance entre la taille des dents et le sexe d’un individu est tout de même considérée légèrement significative (Kieser 1990). En effet, les dents des hommes semblent en moyenne 4% plus grandes avec les canines possédant les divergences les plus marquées et les incisives les moins évidentes (Garn et al. 1964). Ainsi, at the simplest level, differences in tooth size between male and female humans reside mainly in the canines (Kieser 1990, p. 67). Une seule variable appartenant au district des canines se révèle souvent plus prononcée avec des incidences plus élevées chez l’homme et représente, ainsi, le trait le plus sexuellement dimorphique. Il s’agit de la crête accessoire distale (Scott 1975, 1977). C.G. Turner, C.R. Nichol et G.R. Scott (1991) considèrent pour cette variable que observers may not wish to pool sex in populations studies where sex ratios among samples are unequal (Turner et al. 1991, p. 17). Il est donc important de tester si une telle relation existe au sein d’un échantillon. Si la mesure ne peut être réalisée, il faudrait éliminer cette variable des analyses. Nous ne nous attarderons pas sur le dimorphisme sexuel de la taille des dents et la relation entre les dimensions et les variations morphologiques dentaires. Les traits non métriques sont des données qui s’expriment soit par la présence/absence soit par une échelle de gradation proportionnelle à la dimension de la dent elle-même. Ces deux points permettent donc de réduire fortement le biais induit par la taille des dents (Scott 1973). Pourquoi s’intéresse-t-on autant à déterminer l’existence ou non d’un dimorphisme sexuel dans l’expression des variations non métriques ? 149 Chapitre 6 La raison principale est que les chercheurs sont souvent amenés à travailler sur des fréquences populationnelles sexes confondus. Certains, comme J.T. Mayhall (1992), pensent que les différences sexuelles dans l’expression d’une variable et les incidences sont négligeables, permettant ainsi de combiner les sexes au sein des analyses. D’autres contestent cette position, comme E.F. Harris (1980), ou du moins, considèrent qu’il faudrait vérifier que l’on puisse exclure tout biais induit par le dimorphisme sexuel des variables. Quoi qu’il en soit, selon les spécificités des échantillons, il est parfois impossible de travailler différemment qu’en passant sous silence cet aspect. Dans le meilleur des cas, il est possible de contrôler la représentativité d’un échantillon. Si le recrutement funéraire est équilibré, les incidences du dimorphisme sexuel sont minimes. En revanche, si la représentativité ne l’est pas, l’intégration de variables influencées par le sexe des individus pourrait conduire à des interprétations biaisées par la mauvaise composition des échantillons. Un certain nombre d’études ne prennent ou ne peuvent pas prendre en considération le rôle du dimorphisme sexuel dans l’expression des traits non métriques dentaires (Coppa et al. 1995, 1998, Eades 2003...). D’autres testent l’influence du sexe des individus afin d’éliminer les variables qui se révèleraient corrélées pour ne pas introduire de biais de ce type dans les analyses (Johnson et Lovell 1995, Irish 1998, Hsu et al. 1999...). 6.4.1. Le test de la dépendance du sexe dans l’expression des traits Il ne semble pas y avoir de règle pré-établie concernant la procédure à suivre. De ce fait, nous décidons de réaliser une vérification de la présence ou de l’absence d’un dimorphisme sexuel pour les traits dentaires intégrés dans ce travail. Ceci peut être utile puisque nous ne pouvons pas contrôler la représentativité des populations étudiées. Nous aurions souhaité pouvoir examiner cet aspect pour chaque population. Malheureusement, cette mesure ne peut être effectuée sur l’ensemble des échantillons. Des déterminations sexuelles sont disponibles pour deux régions seulement : la Bohême et la Suisse. En effet, pour la Hongrie, les individus étudiés ne sont pas sexés et pour le sud de la France et le nord de l’Espagne, la très large majorité des échantillons sont composés de dents isolées. De plus, quelques considérations sont nécessaires: • Les effectifs mobilisables des régions pour lesquelles cette mesure peut être réalisée représentent une partie limitée des échantillons (251 individus sur plus de 500 pour la Bohême et 116 individus sur plus de 600 pour la Suisse). • Les méthodes ayant servi à déterminer le sexe des individus ne sont pas toujours connues. Nous ne reviendrons pas ici sur les difficultés de la fiabilité rencontrées par certaines de ces méthodes, néanmoins, il faut en avoir conscience et rester prudent. Nous décidons, malgré toutes les difficultés énoncées, d’effectuer cette analyse, tout en gardant un œil attentif aux interprétations qui peuvent découler des résultats que nous obtiendrons. Nous employons le test du Chi2, méthode statistique non paramétrique, pour mettre en évidence d’éventuels caractères tributaires d’une influence sexuelle. Nous déterminons les hypothèses de départ suivantes : H0 = hypothèse nulle : l’expression d’un trait est dépendant du sexe d’un individu. H1 = hypothèse alternative : l’expression d’un trait est indépendant du sexe d’un individu. Les hypothèses sont testées au seuil de signification de 0.05 et de 0.01. Selon le seuil de signification retenu, un trait est significativement lié au sexe d’un individu si la valeur obtenue lui est inférieure. Les calculs sont réalisés sur l’ensemble des variables (n=164) observées sur la dentition permanente. 150 Le traitement préliminaire des données 6.4.1.1. Les résultats concernant le dimorphisme sexuel des traits pour la Bohême Pour la Bohême, seuls 251 individus possèdent une attribution sexuelle sur l’ensemble de l’échantillon qui se compose de 507 individus. Sur les 164 traits de départ, 119 caractères (73%) ont fait l’objet d’un calcul (figure 61). Les résultats de cette mesure se trouvent en table 27. La différence entre les sexes n’est pas significative pour 115 variables (97%) ayant fait l’objet de cette mesure. Seuls quatre traits présentent des divergences significatives à un seuil de 0.05. Il s’agit : - de l’hypocone (T26) observé sur la deuxième molaire supérieure (p=0.049). - de l’hypocone (T26) observé sur la première molaire supérieure (p=0.034). - des crêtes linguales marginales (T43) observées sur l’incisive latérale inférieure (p=0.035). - de l’hypoconulide-entoconulide (T55-56) observé sur la première molaire inférieure (p=0.042). DENTS PERMANENTES Maxillaire 1 Absence congénitale 2 Nombre de radicules 3 Racine appendiciforme 4 Nombre de racines des prémolaires 5 Nombre de racines des molaires 6 Racine de Carabelli 7 Racine paramolaire 8 Racine idiopathique 9 Degré de rotation 10 Convexité vestibulaire 11 Sillon traversant 12 Tubercule distal 13 Cingulum vestibulaire 14 Gouttière vestibulaire 15 Crêtes linguales marginales 16 Tubercule dentaire 17 Crêtes vestibulaires marginales 18 Crête mésiale 19 Crête accessoire distale 20 Cuspides accessoires 21 Odontome 22 Extension de l'émail 23 Parastyle 24 Trait de Carabelli 25 Métacone 26 Hypocone 27 Métaconule 28 Tubercule distal accessoire 29 Tubercule mésial paracone 30 Tubercule mésial accessoire 31 Protoconule 32 Tubercule lingual paracone M3 M2 M1 P2 P1 C I2 I1 corrélation significative pas de corrélation ● non calculé ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● Mandibule ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● 33 Absence congénitale 34 Nombre de radicules 35 Racine appendiciforme 36 Nombre racines canines 37 Racine de Tomes 38 Nombre racines molaires 39 Racine de Citroen 40 Racine paramolaire 41 Racine entomolaire 42 Racine idiopathique 43 Crêtes linguales marginales 44 Crêtes vestibulaires marginales 45 Crête accessoire distale 46 Odontome 47 Variation des cuspides linguales 48 Pli cuspide mésiolinguale 49 Fosse précuspidale 50 Crête trigonide intermédiaire 51 Crête trigonide distale 52 Nombre de cuspides 53 Dessin du sillon 54 Protostylide 55-56 Hypoconulide-Entoconulide 57 Métaconulide 58 Tubercule paracone 59 Tubercule de Citroen M3 M2 M1 P2 P1 C I2 I1 ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● Figure 61 : Représentation synthétique touchant les résultats du test de la dépendance du sexe dans l’expression des traits non métriques dentaires pour la Bohême. 6.4.1.2. Les résultats concernant le dimorphisme sexuel des traits pour la Suisse Pour la Suisse, seuls 116 individus possèdent une attribution sexuelle sur l’ensemble de l’échantillon qui se compose de plus de 620 individus. Sur les 164 traits de départ, 109 caractères (67%) ont fait l’objet d’un calcul (figure 62). Les résultats de cette mesure se trouvent en table 28. La différence entre les sexes n’est pas significative pour 109 variables (94%) ayant fait l’objet de cette mesure. Sept traits présentent des divergences significatives à un seuil de 0.05. Il s’agit : - du nombre de radicules (T2) observé sur la troisième molaire supérieure (p=0.03). - de l’extension de l’émail (T22) observée sur la troisième molaire supérieure (p=0.039). 151 Chapitre 6 - du métaconulide (T27) observé sur la troisième molaire supérieure (p=0.032) - du nombre de radicules (T34) observé sur la deuxième molaire inférieure (p=0.02) - du nombre de radicules (T34) observé sur l’incisive centrale inférieure (p=0.034) - de la crête trigonide intermédiaire (T50) observée sur la deuxième molaire inférieure (p=0.02) - du dessin du sillon (T53) sur la troisième molaire inférieure (p=0.039) DENTS PERMANENTES Maxillaire 1 Absence congénitale 2 Nombre de radicules 3 Racine appendiciforme 4 Nombre de racines des prémolaires 5 Nombre de racines des molaires 6 Racine de Carabelli 7 Racine paramolaire 8 Racine idiopathique 9 Degré de rotation 10 Convexité vestibulaire 11 Sillon traversant 12 Tubercule distal 13 Cingulum vestibulaire 14 Gouttière vestibulaire 15 Crêtes linguales marginales 16 Tubercule dentaire 17 Crêtes vestibulaires marginales 18 Crête mésiale 19 Crête accessoire distale 20 Cuspides accessoires 21 Odontome 22 Extension de l'émail 23 Parastyle 24 Trait de Carabelli 25 Métacone 26 Hypocone 27 Métaconule 28 Tubercule distal accessoire 29 Tubercule mésial paracone 30 Tubercule mésial accessoire 31 Protoconule 32 Tubercule lingual paracone M3 M2 M1 P2 P1 ● C I2 corrélation significative pas de corrélation ● non calculé I1 ● ● ● ● ● ● ● ● ● ● ● ● ● Mandibule ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● 33 Absence congénitale 34 Nombre de radicules 35 Racine appendiciforme 36 Nombre racines canines 37 Racine de Tomes 38 Nombre racines molaires 39 Racine de Citroen 40 Racine paramolaire 41 Racine entomolaire 42 Racine idiopathique 43 Crêtes linguales marginales 44 Crêtes vestibulaires marginales 45 Crête accessoire distale 46 Odontome 47 Variation des cuspides linguales 48 Pli cuspide mésiolinguale 49 Fosse précuspidale 50 Crête trigonide intermédiaire 51 Crête trigonide distale 52 Nombre de cuspides 53 Dessin du sillon 54 Protostylide 55-56 Hypoconulide-Entoconulide 57 Métaconulide 58 Tubercule paracone 59 Tubercule de Citroen M3 M2 M1 P2 P1 C I2 I1 ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● Figure 62 : Représentation synthétique touchant les résultats du test de la dépendance du sexe dans l’expression des traits non métriques dentaires pour la Suisse. 6.4.1.3. Les résultats concernant le dimorphisme sexuel des traits : un bilan La décision concernant les manipulations se révèle ardue à l’issue des résultats de la mesure de la dépendance du sexe dans l’expression des traits non métriques dentaires. Différents aspects étaient connus au préalable annonçant la difficulté de cette manœuvre. En effet, l’analyse a été possible sur deux régions seulement présentant des effectifs en détermination sexuelle limités. De plus, nous avons vu que pour défendre l’idée d’une signification biologique, les différences dans l’expression d’une variable devraient présenter une aptitude interpopulationnelle. Tel n’a pas été le cas. En effet, chaque région possède une série de variables différentes, - quatre pour la Bohême et sept pour la Suisse -, présentant des divergences statistiques significatives entre les sexes des individus, toutes à un seuil d’acceptation de 0.05. Ces éléments, - la non-persistance des corrélations et les seuils d’acceptation -, nous induisent à considérer que les différences qui se sont révélées statistiquement significatives sont probablement dues à des biais d’échantillonnage. Ainsi, nous décidons de conserver l’hypothèse 1 avec le risque de commettre une erreur de type I pour toutes les variables (conclure de manière erronée que les expressions des traits non métriques sont indépendantes du sexe des individus). 152 Le traitement préliminaire des données 6.5. L’essentiel en bref Ce chapitre a abordé le traitement préliminaire des observations dentaires. Il a comporté divers aspects, telles la vérification, l’épuration et l’optimisation des données en vue des analyses. Il s’agit d’une phase de manipulations des données délicate, impliquant des décisions importantes. Elle s’articule de la manière suivante : • Statistiques descriptives Cette partie a concerné une phase descriptive des données brutes. Nous avons, dans un premier temps, numérisé les variables alphabétiques afin de pouvoir les traiter statistiquement (T11 et T53 sur la dentition permanente). Nous avons, dans un deuxième temps, réalisé un décompte des observations par dentition pour chaque région. Près de 255'000 données sur plus de 2'000 individus ont été enregistrées pour cette étude. Nous avons, ensuite, procédé à une étape de vérification de la transcription des observations sur support informatique (erreurs de frappe, gradations aberrantes, décalage de saisie...) afin d’éliminer le cas échéant les erreurs ayant pu s’y glisser. Enfin, nous avons évalué la pertinence des variables observées. Les traits ne présentant pas un effectif suffisant ont été éliminés. La suppression concerne uniquement les variables observées sur la dentition déciduale pour la Bohême, la Hongrie et le nord de l’Espagne. Les données constantes n’ont pas été éliminées ici. • Asymétrie des antimères Cette étape était essentielle puisqu’elle a touché la transformation des observations bilatérales en données uniques afin d’éliminer les informations redondantes fournies par les antimères. Elle a d’abord consisté à mesurer le degré de similarité des expressions des traits dentaires observés sur les dents controlatérales, et ce, pour chacune des régions. Nous avons vu que les relations et l’intensité des expressions antimériques dépendent de la spécificité des échantillons étudiés. Ainsi, les meilleurs résultats ont été obtenus pour la Suisse et la Bohême, constituées en majorité de sujets individualisés. Les traits non corrélés variant d’une région à l’autre, nous avons choisi de considérer les expressions antimériques de l’ensemble des variables interdépendantes. Ensuite, le traitement des données bilatérales a été réalisé. Parmi les différentes méthodes employées, deux d’entre elles ont été retenues : le comptage latéral - qui consiste à ne retenir qu’un antimère (nous avons choisi arbitrairement le gauche) - et le comptage individuel - qui vise à ne garder qu’une expression et de préférence la plus élevée -. La sélection de l’une ou l’autre de ces méthodes a été opérée en deux temps distincts : 1) En fonction du type de variables : six traits - qui pour la plupart traduisent une forme ou une position ont fait l’objet d’un traitement particulier. Le comptage individuel est réalisé pour T9, T11 (après dichotomisation) et T55-56 (après fusion de ces deux traits et une nouvelle gradation) observés sur la dentition permanente. Le comptage latéral est sélectionné pour T53 sur la dentition permanente, T3, T11 et T36 sur la dentition déciduale. Ces décisions s’appliquent sur l’ensemble des populations étudiées. 2) En fonction de l’état des échantillons : la Bohême et la Hongrie font l’objet d’un traitement par comptage individuel. Les trois autres régions - présentant des situations composites - ont été manipulées selon la spécificité des sites : le comptage individuel pour les sites consentant une individualisation des sujets et le comptage latéral pour ceux offrant une majorité de dents isolées. • Corrélations entre les traits Cette fraction a concerné l’évaluation des relations entre les variables pour l’ensemble des populations. Nous nous sommes intéressée à trois types de liens : 1) Les corrélations intradistrict : nous avons vérifié que les variables enregistrées suivent le modèle attendu nommé gradient d’expression ou du champ (si un trait est présent sur un membre d’une classe dentaire, il est fort probable de l’observer sur l’ensemble du groupe avec un accroissement de l’expression mésio-distal). Les résultats montrent que les traits ne suivant pas ce modèle varient selon les 153 Chapitre 6 populations étudiées. Pour des raisons de représentativité d’échantillon, nous choisissons qu’un trait ne se conforme pas au modèle si les résultats de deux régions majeures (CZ et CH) ou une région majeure et au moins deux régions mineures (HU, F, ES) vont dans ce sens. Ainsi, trois variables (T15, T53 et T57) ne suivent pas le gradient d’expression. 2) Les corrélations interdistrict : nous avons testé l’interdépendance d’une variable s’exprimant au-delà d’un district dentaire. L’objectif était de définir si l’effet du champ pouvait également être mis en évidence sur plusieurs classes dentaires. La même procédure de décision a été réalisée et une seule variable (T16) s’est révélée statistiquement significative. 3) Les corrélations intertrait : nous avons mesuré une association entre variables s’exprimant sur les arcades opposées, soit la relation entre le tubercule de Carabelli (T24) observé sur les molaires supérieures et le protostylide (T54) sur les inférieures. Nous l’avons sélectionnée puisqu’il semblerait que ce soit la corrélation la plus souvent rencontrée dans la littérature. Une seule association s’est révélée positive, soit l’expression du tubercule de Carabelli de la troisième molaire supérieure avec l’expression du protostylide de la deuxième molaire inférieure. • Dimorphisme sexuel Ce dernier point a touché la mesure de l’influence du sexe dans l’expression des traits non métriques dentaires. L’analyse a été réalisée sur deux régions seulement (CZ et CH) composées par des effectifs limités en détermination sexuelle. Pour prétendre qu’une signification biologique est présente, les différences dans l’expression d’une variable devraient présenter une aptitude interpopulationnelle. Ainsi, nous avons choisi de retenir une association comme pertinente seulement si elle se révèle significative pour les deux populations testées. Or, aucune des variables concernées n’a satisfait à cette exigence. Il n’y a pas un traitement préliminaire des données dentaires standard. Selon les données et les résultats des différents tests, nous avons pris certaines décisions. Selon les analyses, il sera peut-être utile et possible de prêter attention à certains résultats du traitement préliminaire afin d’intégrer ou d’exclure certaines variables. 154 Chapitre 7 L’analyse des données Ce chapitre - abordant l’analyse des données - s’articule en deux parties. La première relate la procédure des traitements statistiques préliminaires et analytiques appliquée à toutes les populations. La seconde présente les résultats obtenus pour chaque ensemble - région ou groupe - testé. 7.1. La procédure suivie Un traitement statistique préliminaire des données dentaires est nécessaire avant d’aborder l’analyse. La démarche est la même pour chaque région. Comme nous ne travaillons pas avec des données individuelles mais avec des fréquences populationnelles, la première étape consiste à calculer les incidences des traits au sein des différentes populations. Les variables ne sont pas toutes dichotomiques, c’est pourquoi il faut ensuite trouver le meilleur moyen de garder la variabilité des traits gradués à travers leur fréquence. La phase suivante se traduit par la sélection des variables mobilisées au sein des analyses. Enfin, un aperçu des diverses méthodes d’analyses des données mobilisées est réalisé. Les tables correspondant aux diverses étapes du traitement préliminaire seront désignées lors de la présentation des analyses des différents ensembles. 7.1.1. Le calcul d’incidences populationnelles Nous avons calculé les incidences et réalisé un décompte des observations pour chaque variable. Ces mesures ont été réalisées sur l’ensemble des régions et des groupes testés pour la dentition permanente. Pour celle déciduale, seuls la Bohême, le sud de la France et la Suisse ont fait l’objet d’un calcul. L’exploitation des données provenant de la dentition temporaire sera uniquement descriptive. Leurs effectifs étant largement insuffisants, nous ne pourrons malheureusement pas les intégrer aux phases analytiques. Cette première étape permet d’avoir une vision globale des données fournies par la morphologie dentaire. Elle participe également à la détermination des groupes et des variables qui pourront être intégrés dans les analyses. Les groupes sélectionnés dépendent de la spécificité des échantillons. Ainsi, les ensembles correspondent parfois à des sites, parfois à un regroupement d’individus. L’objectif étant d’obtenir des effectifs suffisants pour les analyses. La constitution des échantillons sera indiquée de cas en cas. Nous avons choisi un seuil minimal de cinq observations pour qu’une variable ou un groupe soit retenu. Selon les cas, nous avons préféré éliminer un ensemble plutôt que des variables. 7.1.2. Le calcul de fréquences ajustées L’intégration des données sous forme de fréquences dans les analyses semble à première vue ne poser aucun problème. Ceci est le cas pour les variables dichotomiques pour lesquelles l’incidence de leur présence est retenue. Pour les variables graduées, plusieurs solutions sont possibles : réduire l’information par la présence ou l’absence du trait (dichotomie) ou par la définition d’un seuil d’expression variant selon le trait considéré (breakpoint) ou encore garder la variabilité de l’expression des traits gradués par l’emploi de fréquences ajustées (expression count). Les deux premières méthodes réduisent l’ensemble de l’information fournie par les gradations. De plus, des travaux portant sur la mesure de l’héritabilité des traits non métriques crâniens ont montré que le 155 Chapitre 7 choix d’un seuil (breakpoint) pour les variables graduées influence de façon considérable les résultats et doit être réalisé avec précaution (Carson 2006). Pour ces différentes raisons, nous avons choisi de retenir la méthode dite de l’expression count mise en place par C.G. Turner (1985). Elle permet d’obtenir un pourcentage de présence unique en ajustant les fréquences d’expression pour chaque stade à l’aide d’un coefficient qui est proportionnel au nombre de stades observés et en sommant ces nouvelles expressions. Nous obtenons ainsi une fréquence unique (F.C. - frequence count) tenant compte de l’information fournie par les gradations. Un facteur de correction prenant en compte la taille de l’échantillon est utilisé. Il s’agit simplement de multiplier la fréquence unique par une valeur correspondant au nombre d’observations réalisées (n) divisé par le nombre d’observations possibles (N). Nous obtenons alors un dénombrement d’expression (E.C. expression count). Prenons, à titre d’exemple, l’absence congénitale de la troisième molaire supérieure (T1M3), qui est une variable présentant trois expressions positives différentes (stade 0 : taille de la dent normale ; stade 1 : réduction en taille de la couronne ; stade 2 : dent en pic ; stade 3 : absence de la dent). La démarche pour obtenir son E.C. - expression count - est la suivante : 1. Le calcul du coefficient Selon le nombre de stades positifs, le coefficient varie d’un trait à l’autre. Celui-ci s’obtient par la division d’une constante égale à 1 par le nombre de stades observés (ici 3 stades positifs : ST3). Dans notre exemple le coefficient est égal à 0.33 (= 1/3). 2. La multiplication du coefficient par les stades observés L’étape suivante consiste à multiplier le coefficient par le stade correspondant, dans notre exemple : - le coefficient stade 0 = 0.33*0 = 0 - le coefficient stade 1 = 0.33*1 = 0.33 - le coefficient stade 2 = 0.33*2 = 0.67 - le coefficient stade 3 = 0.33*3 = 1 3. L’obtention d’une fréquence ajustée La phase suivante comprend la multiplication de chaque fréquence par son coefficient, ici : - la fréquence ajustée/stade 0 = fréquence * coefficient stade 0 = fréq0*0 - la fréquence ajustée/stade 1 = fréquence * coefficient stade 1 = fréq1*0.33 - la fréquence ajustée/stade 2 = fréquence * coefficient stade 2 = fréq2*0.67 - la fréquence ajustée/stade 3 = fréquence * coefficient stade 3 = fréq3*1 4. L’addition des fréquences ajustées : frequence count - F.C. Il s’agit de sommer chaque fréquence ajustée pour obtenir une F.C. F.C. = fréq_ajustées 5. Le calcul de l’expression count - E.C. L’ultime étape introduisant le facteur de correction de la taille de l’échantillon aboutit à l’E.C. E.C. = F.C.* n/N Nous avons appliqué cette méthode à l’ensemble des variables observées sur la dentition permanente. La figure 63 synthétise les formules utilisées pour chaque trait afin d’obtenir des F.C. - frequence count. Figure 63 : Liste des formules utilisées pour chaque variable en vue de la transformation des incidences en fréquences uniques F.C. - frequence count ; avec - dans l’ordre - le nom de la variable, le nombre de stades positifs (Gr.) et la formule employée. 156 L’analyse des données Nom du trait 1 Absence congénitale P2 1 Absence congénitale M3+I2 2 Nombre de radicules 3 Racine appendiciforme 4 Nombre de racines prémolaires 5 Nombre de racines molaires 6 Racine de Carabelli 7 Racine paramolaire 8 Racine idiopathique 9 Degré de rotation 10 Convexité vestibulaire 11 Sillon traversant 12 Tubercule distal 13 Cingulum vestibulaire 14 Gouttière vestibulaire 15 Crêtes linguales marginales C 15 Crêtes linguales marginales I 16 Tubercule dentaire I1 16 Tubercule dentaire I2+C Gr. ST1 ST3 1-4 ST1 ST2 ST3 ST1 ST1 ST1 ST1 ST4 ST1 ST1 ST1 ST1 ST5 ST6 ST4 ST7 17 Crêtes vestibulaires marginales 18 Crête mésiale 19 Crête accessoire distale 20 Cuspides accessoires 21 Odontome 22 Extension de l'émail 23 Parastyle 24 Trait de Carabelli ST3 ST3 ST5 ST1 ST1 ST3 ST6 ST7 25 Métacone 26 Hypocone 27 Métaconule 28 Tubercule distal accessoire 29 Tubercule mésial paracone 30 Tubercule mésial accessoire 31 Protoconule 32 Tubercule lingual paracone 33 Absence congénitale P2 33 Absence congénitale M3+I1 34 Nombre de radicules 35 Racine appendiciforme 36 Nombre racines canines 37 Racine de Tomes 38 Nombre racines molaires 39 Racine de Citroën 40 Racine paramolaire 41 Racine entomolaire 42 Racine idiopathique 43 Crêtes linguales marginales 44 Crêtes vestibulaires marginales 45 Crête accessoire distale 46 Odontome 47 Variation des cuspides linguales ST6 ST6 ST1 ST5 ST1 ST1 ST1 ST1 ST1 ST3 1-5 ST1 ST1 ST5 ST2 ST1 ST1 ST1 ST1 ST3 ST3 ST5 ST1 ST9 48 Pli cuspide mésiolinguale 49 Fosse précuspidale 50 Crête trigonide intermédiaire 51 Crête trigonide distale 52 Nombre de cuspides 53 Dessin du sillon (Y+X) 53 Protostylide ST3 ST4 ST1 ST1 ST2 ST2 ST7 55-56 Hypoconulide-Entoconulide ST8 57 Métaconulide 58 Tubercule paracone 59 Tubercule de Citroën ST5 ST1 ST1 Formule (Fréq0*0)+(Fréq1*1) (Fréq0*0)+(Fréq1*0.33)+(Fréq2*0.67)+(Fréq3*1) variable selon le district (Fréq0*0)+(Fréq1*1) (Fréq1*0)+(Fréq2*0.5)+(Fréq3*1) (Fréq1*0)+(Fréq2*0.33)+(Fréq3*0.67)+(Fréq4*1) (Fréq0*0)+(Fréq1*1) (Fréq0*0)+(Fréq1*1) (Fréq0*0)+(Fréq1*1) (Fréq0*0)+(Fréq1*1) (Fréq0*0)+(Fréq1*0.25)+(Fréq2*0.5)+(Fréq3*0.75)+(Fréq4*1) (Fréq0*0)+(Fréq1*1) (Fréq0*0)+(Fréq1*1) (Fréq0*0)+(Fréq1*1) (Fréq0*0)+(Fréq1*1) (Fréq0*0)+(Fréq1*0.2)+(Fréq2*0.4)+(Fréq3*0.6)+(Fréq4*0.8)+(Fréq5*1) (Fréq0*0)+(Fréq1*0.167)+(Fréq2*0.334)+(Fréq3*0.501)+(Fréq4*0.668)+(Fréq5*0.835)+(Fréq6*1) (Fréq0*0)+(Fréq1*0.25)+(Fréq2*0.5)+(Fréq3*0.75)+(Fréq4*1) (Fréq0*0)+(Fréq1*0.143)+(Fréq2*0.288)+(Fréq3*0.431)+(Fréq4*0.574)+(Fréq5*0.717)+(Fréq6*0.86) +(Fréq7*1) (Fréq0*0)+(Fréq1*0.33)+(Fréq2*0.67)+(Fréq3*1) (Fréq0*0)+(Fréq1*0.33)+(Fréq2*0.67)+(Fréq3*1) (Fréq0*0)+(Fréq1*0.2)+(Fréq2*0.4)+(Fréq3*0.6)+(Fréq4*0.8)+(Fréq5*1) (Fréq0*0)+(Fréq1*1) (Fréq0*0)+(Fréq1*1) (Fréq0*0)+(Fréq1*0.33)+(Fréq2*0.67)+(Fréq3*1) (Fréq0*0)+(Fréq1*0.167)+(Fréq2*0.334)+(Fréq3*0.501)+(Fréq4*0.668)+(Fréq5*0.835)+(Fréq6*1) (Fréq0*0)+(Fréq1*0.143)+(Fréq2*0.288)+(Fréq3*0.431)+(Fréq4*0.574)+(Fréq5*0.717)+(Fréq6*0.86) +(Fréq7*1) (Fréq0*0)+(Fréq1*0.167)+(Fréq2*0.334)+(Fréq3*0.501)+(Fréq4*0.668)+(Fréq5*0.835)+(Fréq6*1) (Fréq0*0)+(Fréq1*0.167)+(Fréq2*0.334)+(Fréq3*0.501)+(Fréq4*0.668)+(Fréq5*0.835)+(Fréq6*1) (Fréq0*0)+(Fréq1*1) (Fréq0*0)+(Fréq1*0.2)+(Fréq2*0.4)+(Fréq3*0.6)+(Fréq4*0.8)+(Fréq5*1) (Fréq0*0)+(Fréq1*1) (Fréq0*0)+(Fréq1*1) (Fréq0*0)+(Fréq1*1) (Fréq0*0)+(Fréq1*1) (Fréq0*0)+(Fréq1*1) (Fréq0*0)+(Fréq1*0.33)+(Fréq2*0.67)+(Fréq3*1) variable selon le district (Fréq0*0)+(Fréq1*1) (Fréq1*0)+(Fréq2*1) (Fréq0*0)+(Fréq1*0.2)+(Fréq2*0.4)+(Fréq3*0.6)+(Fréq4*0.8)+(Fréq5*1) (Fréq1*0)+(Fréq2*0.5)+(Fréq3*1) (Fréq0*0)+(Fréq1*1) (Fréq0*0)+(Fréq1*1) (Fréq0*0)+(Fréq1*1) (Fréq0*0)+(Fréq1*1) (Fréq0*0)+(Fréq1*0.33)+(Fréq2*0.67)+(Fréq3*1) (Fréq0*0)+(Fréq1*0.33)+(Fréq2*0.67)+(Fréq3*1) (Fréq0*0)+(Fréq1*0.2)+(Fréq2*0.4)+(Fréq3*0.6)+(Fréq4*0.8)+(Fréq5*1) (Fréq0*0)+(Fréq1*1) (Fréq0*0)+(Fréq1*0.11)+(Fréq2*0.22)+(Fréq3*0.33)+(Fréq4*0.44)+(Fréq5*0.55)+(Fréq6*0.66) +(Fréq7*0.77)+ (Fréq8*0.88)+(Fréq9*1) (Fréq0*0)+(Fréq1*0.33)+(Fréq2*0.67)+(Fréq3*1) (Fréq0*0)+(Fréq1*0.25)+(Fréq2*0.5)+(Fréq3*0.75)+(Fréq4*1) (Fréq0*0)+(Fréq1*1) (Fréq0*0)+(Fréq1*1) (Fréq4*0)+(Fréq5*0.5)+(Fréq6*1) (Fréq1*0)+(Fréq2*0.5)+(Fréq3*1) (Fréq0*0)+(Fréq1*0.143)+(Fréq2*0.288)+(Fréq3*0.431)+(Fréq4*0.574)+(Fréq5*0.717)+(Fréq6*0.86) +(Fréq7*1) (Fréq0*0)+(Fréq1*0.125)+(Fréq2*0.25)+(Fréq3*0.375)+(Fréq4*0.5)+(Fréq5*625)+(Fréq6*0.75) +(Fréq7*0.875)+(Fréq8*1) (Fréq0*0)+(Fréq1*0.2)+(Fréq2*0.4)+(Fréq3*0.6)+(Fréq4*0.8)+(Fréq5*1) (Fréq0*0)+(Fréq1*1) (Fréq0*0)+(Fréq1*1) 157 Chapitre 7 7.1.3. La sélection des variables intégrées dans les analyses Il n’y a pas de procédure standard de sélection des variables mobilisables au sein des analyses. L’ensemble de données non fusionnées ou encore une partie seulement constituée des variables observées sur les dents dites clés (key tooth) peuvent être intégrées dans la phase analytique. Nous avons vu qu’une très large majorité de variables s’exprimant sur les dents controlatérales, ainsi qu’à l’intérieur d’une même classe dentaire (sauf T15, T53 et T57), ne sont pas indépendantes les unes des autres. Un trait montre également une interdépendance interdistrict (T16). Aucun dimorphisme sexuel n’a été retenu comme significatif lors du traitement préliminaire des données. Nous avons choisi d’éviter d’introduire la redondance d’informations, c’est pourquoi seuls les traits s’exprimant sur une dent par district sont retenus. Il s’agit généralement du membre mésial de la classe dentaire. Nous avons sélectionné les dents clés suivant les conseils de C.R. Scott et C.G. Turner (1997), soit l’incisive centrale supérieure, l’incisive latérale inférieure, les canines et les premières molaires. Pour les prémolaires, nous avons considéré la partie de la dent montrant une variabilité plus accentuée. Ainsi, nous avons gardé les traits observés sur la première prémolaire supérieure, sur la racine de la première prémolaire inférieure et la couronne de la seconde. Pour les trois variables qui ne semblent pas suivre le gradient d’expression, nous choisissons de retenir uniquement la dent clé (T15-I1, T53-M1 et T57-M1). Pour le tubercule dentaire (T16), variable significativement liée interdistrict, nous incluons, selon les populations testées, l’une des deux dents clés seulement (I1 ou C). Nous avons également gardé l’ensemble des membres dentaires sur lesquels il est possible d’enregistrer leur absence congénitale (T1). Ainsi, les traits potentiellement mobilisables dans la phase analytique s’élèvent à 79 observations et sont représentés par un point dans la figure 64. DENTS PERMANENTES Maxillaire 1 Absence congénitale 2 Nombre de radicules 3 Racine appendiciforme 4 Nombre de racines des prémolaires 5 Nombre de racines des molaires 6 Racine de Carabelli 7 Racine paramolaire 8 Racine idiopathique 9 Degré de rotation 10 Convexité vestibulaire 11 Sillon traversant 12 Tubercule distal 13 Cingulum vestibulaire 14 Gouttière vestibulaire 15 Crêtes linguales marginales 16 Tubercule dentaire 17 Crêtes vestibulaires marginales 18 Crête mésiale 19 Crête accessoire distale 20 Cuspides accessoires 21 Odontome 22 Extension de l'émail 23 Parastyle 24 Trait de Carabelli 25 Métacone 26 Hypocone 27 Métaconule 28 Tubercule distal accessoire 29 Tubercule mésial paracone 30 Tubercule mésial accessoire 31 Protoconule 32 Tubercule lingual paracone M3 M2 M1 P2 P1 ● C ● ● I2 I1 ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● Mandibule ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● 33 Absence congénitale 34 Nombre de radicules 35 Racine appendiciforme 36 Nombre racines canines 37 Racine de Tomes 38 Nombre racines molaires 39 Racine de Citroen 40 Racine paramolaire 41 Racine entomolaire 42 Racine idiopathique 43 Crêtes linguales marginales 44 Crêtes vestibulaires marginales 45 Crête accessoire distale 46 Odontome 47 Variation des cuspides linguales 48 Pli cuspide mésiolinguale 49 Fosse précuspidale 50 Crête trigonide intermédiaire 51 Crête trigonide distale 52 Nombre de cuspides 53 Dessin du sillon 54 Protostylide 55-56 Hypoconulide-Entoconulide 57 Métaconulide 58 Tubercule paracone 59 Tubercule de Citroen M3 M2 M1 P2 P1 ● C I2 ● ● I1 ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● Figure 64 : Schéma synthétisant la sélection des traits observés sur les dents clés qui seront intégrés dans la phase analytique. 158 L’analyse des données 7.1.4. Les analyses mobilisées La phase analytique comporte l’intégration de plusieurs méthodes multivariées complémentaires. Les analyses sont effectuées à partir de fréquences ajustées par la méthode de l’E.C. - expression count. Nous avons choisi deux types de représentations des distances entre échantillons : l’arbre et le nuage de points. Toutes les analyses sont effectuées à l’aide du logiciel PAST - PAlaeontological STatistics, version 1.67 (Hammer et Harper 2005). Configuration sous forme d’arbre • L’analyse de classification ascendante hiérarchique selon la méthode de Ward (hierarchical cluster analysis - Ward method) Cette méthode de classification assemble les données en clusters en employant une mesure de distance entre les différents échantillons étudiés. Nous avons choisi l’algorithme d'agrégation de Ward minimisation de la variance intragroupe - (Ward 1963) et la distance euclidienne comme mesure de distance entre les groupes. Les résultats sont proposés sous forme de dendrogrammes. L’analyse du bootstrap Un contrôle de la robustesse des arbres est réalisé par l’analyse du bootstrap développée par B. Efron (1979). Il s’agit d’une méthode de ré-échantillonnage qui consiste à pondérer les caractères de façon aléatoire, dans le but d’évaluer la robustesse d’un arbre (Darlu et Tassy 1993). Cette méthode tire, au hasard avec remise, un ensemble de x caractères et cette procédure peut être réalisée N fois (ici 1000 fois). Ensuite, un dénombrement des associations retrouvées sur l’arbre obtenu par l’analyse ascendante hiérarchique est effectué. Les résultats sont donnés sous forme de pourcentages sur le dendrogramme. Si un regroupement est rencontré à 90%, cela indique que l’on a 10 chances sur 100 de se tromper en affirmant que cette association n’existe pas (Darlu et Tassy 1993). Configuration sous forme de nuage de points • L’analyse des proximités (MDS - multidimensional scaling) Cette méthode réalise une représentation multidimensionnelle des relations entre les individus sur la base de distances par assignation de coordonnées dans l’espace. Elle réorganise les objets de la meilleure manière afin d’arriver à une configuration qui s’approche le plus possible des distances observées. De nouvelles distances, nommées disparités, sont calculées. Une statistique, appelée stress, mesure l’ajustement des distances de départ aux disparités calculées. Plus elle est petite, plus l’adéquation est bonne (entre 0 et 5 %). Les résultats se présentent sous forme de nuages de points. Cette méthode est considérée comme étant une alternative aux analyses factorielles, mais contrairement à celles-ci, elle n’impose pas de restrictions concernant la normalité des données (Dupanloup de Ceuninck 1999). L’arbre minimum couvrant (MST - minimal spanning tree) Nous complétons les représentations sous forme de nuages de points de l’analyse des proximités par un arbre minimum couvrant (MST - minimal spanning tree) pour donner une direction aux relations entre les groupes. Le MST relie les différents groupes à partir des distances euclidiennes calculées sur les données initiales. • L’analyse en composantes principales (ACP) L’analyse en composantes principales est une méthode produisant des variables hypothétiques que l’on nomme composantes. Ces nouvelles variables sont des combinaisons linéaires issues des variables initiales. Cette méthode réduit l’ensemble des données à un nombre réduit de composantes (les plus importantes) afin d’obtenir une image en deux dimensions des relations entre les populations. Les résultats sont donnés sous forme de nuages de points, chaque axe est accompagné d’un indice indiquant la part de la variabilité des données de départ dans la représentation. Nous effectuons cette analyse pour contrôler la robustesse de la configuration proposée par le multidimensional scaling. 159 Chapitre 7 7.2. L’analyse des ensembles Nous entamons ici l’analyse des différents ensembles en abordant plusieurs aspects. Nous débutons par l’analyse des régions appartenant au domaine oriental - la Bohême et la Hongrie -. Elle est, dans un premier temps, intrarégionale. Nous effectuons, dans un second temps, une analyse de l’ensemble de l’aire orientale. Ensuite, nous nous attachons à la situation des régions du domaine méridional, - le sud de la France et le nord de l’Espagne -. La même démarche est effectuée par une première analyse intrarégionale puis élargie à l’aire méridionale. Nous poursuivons par la Suisse, territoire à l’articulation entre les deux domaines. Nous définissons également les rapports de cette région avec les différents domaines étudiés. Les ensembles campaniformes - toutes régions confondues - font également l’objet d’une analyse. La Bohême est la seule région pour laquelle nous avons des déterminations sexuelles suffisantes ; il est intéressant de clore la phase analytique par une étude selon le sexe des individus. Une planche mobile avec un rappel des informations utiles concernant les échantillons est disponible en fin de volume. 7.2.1. Le domaine oriental 7.2.1.1. La Bohême La Bohême symbolise l’occupation orientale type. Elle est généralement considérée comme une région importante dans la compréhension de l’émergence de la culture Campaniforme. Nous étudions pour cette région son substrat, - en partie contemporain -, le Cordé, et la période qui lui succède, la culture d’Unetice (Bronze ancien). 7.2.1.1.1. La constitution des groupes testés Nous avons un échantillon constitué de plus de 500 individus et 64 sites provenant des parties septentrionale et centrale de la Bohême (figure 65). Nous avons décidé de garder les gisements comportant plus de 20 individus et de regrouper les autres sites selon leur localisation et leur attribution culturelle. Le site de Zeleice (n°64) a été éliminé car il s’agit d’un gisement situé en Moravie. Ainsi, nous avons constitué 13 ensembles : 4 pour le Cordé, 4 pour le Campaniforme et 5 pour la culture d’Unetice. Ils se répartissent de la manière suivante : - les ensembles retenus pour le Cordé - les ensembles retenus pour le Campaniforme Vikletice (90 individus), site n° 60 : VIK Bohême occidentale (20 individus), sites n° 18, 20, 42, 43, 44, 46, 48, 53, 62 : BWca Bohême occidentale (58 individus), sites n° 1, 2, 8, 9, 10, 12, 14, 17, 19, 26, 27, 28, 40, 41, 42, 47, 49, 50, 51, 53, 55, 63 : BWco 160 Figure 65 : La Bohême, rappel de la localisation des sites (c.f. figure 9). Bohême orientale (17 individus), sites n° 5, 7, 21, 39 : BEca Bohême orientale (36 individus), sites n° 5, 7 : BEco Bohême centrale (40 individus), sites n° 3, 16, 32, 36, 56, 59 : BCca Bohême centrale (24 individus), sites n° 4, 11, 15, 23, 29, 35, 37, 52, 54 : BCco Mochov (21 individus), site n° 25 : MOC L’analyse des données - les ensembles retenus pour la culture d’Unetice Bohême centrale (36 individus), sites n° 23, 31, 33, 34, 35, 37, 38, 57 : BCun Bohême occidentale (38 individus), sites n° 2, 8, 22, 44, 45, 53, 58, 60, 61: BWun Mala Ohrada (47 individus), site n° 30 : MaOr Brezno (38 individus), site n° 5 : BRE Bohême orientale (41 individus), sites n° 6, 13, 24, 39 : BEun Toutes les observations sur les dents déciduales ont été regroupées selon la période à laquelle elles appartiennent : Cordé, Campaniforme et Unetice. 7.2.1.1.2. Les incidences populationnelles Les incidences et les décomptes pour chaque variable se trouvent en table 29 pour la dentition permanente et en table 30 pour la dentition déciduale. Les ensembles définis pour la dentition définitive possèdent, pour la plupart, suffisamment d’observations pour être intégrés dans les analyses. Aucun de ces groupes n’a été éliminé, et lorsque les données sont insuffisantes pour un ensemble, nous éliminons de préférence la variable. Pour la dentition permanente, les traits observés sur l’ensemble des membres d’un district montrent, en général, des fréquences d’expression très proches les unes des autres (table 29). Les traits observés sur la dentition déciduale, malgré des effectifs fragiles, possèdent des fréquences similaires pour les trois périodes (Cordé, Campaniforme et Unetice) (table 30). Les traits observés sur les racines sont généralement peu variables, ainsi que ceux enregistrés sur les incisives et les canines. Seules les secondes molaires supérieures présentent quelques variations qui ne révèlent aucune différence entre les cultures étudiées. Les fréquences ajustées par la méthode de l’expression count E.C. pour chaque trait observé sur la dentition permanente sont données en table 31. DENTS PERMANENTES Maxillaire 1 Absence congénitale 2 Nombre de radicules 3 Racine appendiciforme 4 Nombre de racines des prémolaires 5 Nombre de racines des molaires 6 Racine de Carabelli 7 Racine paramolaire 8 Racine idiopathique 9 Degré de rotation 10 Convexité vestibulaire 11 Sillon traversant 12 Tubercule distal 13 Cingulum vestibulaire 14 Gouttière vestibulaire 15 Crêtes linguales marginales 16 Tubercule dentaire 17 Crêtes vestibulaires marginales 18 Crête mésiale 19 Crête accessoire distale 20 Cuspides accessoires 21 Odontome 22 Extension de l'émail 23 Parastyle 24 Trait de Carabelli 25 Métacone 26 Hypocone 27 Métaconule 28 Tubercule distal accessoire 29 Tubercule mésial paracone 30 Tubercule mésial accessoire 31 Protoconule 32 Tubercule lingual paracone M3 M2 M1 P2 P1 C ● I2 I1 ● ● ● ● ● ● ● ● Mandibule ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● 33 Absence congénitale 34 Nombre de radicules 35 Racine appendiciforme 36 Nombre racines canines 37 Racine de Tomes 38 Nombre racines molaires 39 Racine de Citroen 40 Racine paramolaire 41 Racine entomolaire 42 Racine idiopathique 43 Crêtes linguales marginales 44 Crêtes vestibulaires marginales 45 Crête accessoire distale 46 Odontome 47 Variation des cuspides linguales 48 Pli cuspide mésiolinguale 49 Fosse précuspidale 50 Crête trigonide intermédiaire 51 Crête trigonide distale 52 Nombre de cuspides 53 Dessin du sillon 54 Protostylide 55-56 Hypoconulide-Entoconulide 57 Métaconulide 58 Tubercule paracone 59 Tubercule de Citroen M3 M2 M1 P2 P1 C I2 ● I1 ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● Figure 66 : Représentation synthétique des dents clés retenues pour les analyses de la Bohême (n=54). 161 Chapitre 7 7.2.1.1.3. Les variables mobilisées dans les analyses Pour la Bohême, 54 variables sur les 79 traits localisés sur les dents clés ont été retenues. 34 d’entre elles s’observent sur le maxillaire et 20 sur la mandibule (figure 66). Les fréquences E.C. qui sont utilisées dans les analyses sont données en table 32. Les traits localisés sur les couronnes ainsi que sur les incisives, les canines et les molaires sont, en général, mieux représentés. Les profils d’expression des variables observées sur les dents clés mobilisables montrent dans les grandes lignes des tendances similaires (figure 67). Il est tout de même possible de distinguer des spécificités selon les groupes. Les profils appartenant à la culture d’Unetice présentent des profils analogues pour les cinq ensembles. Ils se distinguent par la fréquence moins élevée de deux traits (métacone - T25 - et hypocone - T26 -) par rapport à celle rencontrée pour les autres groupes. Les profils des ensembles cordés et campaniformes sont, quant à eux, moins uniformes. Des affinités se discernent entre les Campaniformes de la Bohême occidentale (BEca) avec les Cordés de la même aire géographique (BWco) et du centre de la région (BCco). En revanche, les Cordés de la partie orientale (BEco) s’approchent plus volontiers des ensembles du Bronze ancien. 0.00 T1I2 T1I2 162 0.00 0.10 0.20 0.30 VIK VIK 0.40 0.50 0.60 0.70 T3C T2I1 T2I1 T8M1 T17I1 T17I1 T3C T20P1 T20P1 T8M1 T22P1 T22P1 T17C T23M1 T23M1 T17P1 T27M1 T27M1 T17P1 T29M1 T29M1 T17C T18C T59M1 T33I2 T33I2 T59M1 T57M1 T57M1 T18C T36C T28M1 T28M1 T51M1 T36C T51M1 T22M1 T22M1 T45C T50M1 T50M1 T58M1 T30M1 T30M1 T58M1 T32M1 T32M1 T45C T15C T1M3 T1M3 T54M1 T54M1 T15C T2C T10I1 T10I1 T19C T15I1 T2C T24M1 T19C T15I1 T16C T33M3 T16C T24M1 T11I1 T31M1 T11I1 T31M1 T33M3 T37P1 T53M1 T37P1 T53M1 T4P1 T43I2 T13I1 T13I1 T43I2 T47P2 T47P2 T4P1 T2P1 T34I2 T34I2 T34M1 T34M1 T2P1 T2M1 T52M1 T52M1 T55-56M1 T55-56M1 T2M1 T5M1 T38M1 T38M1 T25M1 T25M1 T5M1 T26M1 T26M1 0.10 0.20 0.30 BWco 0.40 BWco 0.50 0.60 0.70 Profils d’expression des différents groupes définis pour la Bohême 0.10 0.20 0.30 0.40 0.50 0.60 0.70 0.10 0.20 0.30 0.40 0.50 0.60 0.70 T22M1 T45C T10I1 T54M1 T15C T1M3 T32M1 T30M1 T50M1 T22M1 T45C T58M1 T10I1 T54M1 T15C T1M3 T32M1 T30M1 T50M1 T22M1 T45C T58M1 T51M1 T33I2 T18C T59M1 T29M1 T27M1 T23M1 T22P1 T20P1 T17I1 T17C T17P1 T8M1 T3C T2I1 T1I2 T33I2 T18C T59M1 T29M1 T27M1 T23M1 T22P1 T20P1 T17I1 T17C T17P1 T8M1 T3C T2I1 T1I2 0.00 T57M1 T57M1 0.00 T36C T36C T28M1 T36C T28M1 0.00 T1I2 T2I1 T3C T8M1 T17P1 T17C T17I1 T20P1 T22P1 T23M1 T27M1 T29M1 T59M1 T18C T33I2 T57M1 T28M1 T51M1 T51M1 T58M1 T50M1 T30M1 T32M1 T1M3 T15C T54M1 T10I1 T2C T15I1 T2C T2C T19C T24M1 T19C T15I1 T19C T24M1 T33M3 T11I1 T15I1 T24M1 T16C T16C T33M3 T16C T33M3 T31M1 T11I1 T31M1 T11I1 T31M1 T37P1 T53M1 T37P1 T43I2 T53M1 T43I2 T43I2 T4P1 T13I1 T37P1 T4P1 T4P1 T47P2 T34M1 T52M1 T2M1 T55-56M1 T53M1 T13I1 T13I1 T34I2 T2P1 T2P1 T34I2 T34M1 T34M1 T47P2 T52M1 T52M1 T34I2 T2M1 T2M1 T47P2 T2P1 T55-56M1 T55-56M1 T5M1 T38M1 T5M1 T25M1 T26M1 T38M1 0.79 0.83 T5M1 T25M1 T25M1 BCco BCco T38M1 T26M1 T26M1 BEco BEco 0.10 0.20 0.30 0.40 BWca BWca 0.50 0.60 0.70 0.00 T1I2 T2I1 T3C T8M1 T17P1 T17C T17I1 T20P1 T22P1 T23M1 T27M1 T29M1 T59M1 T18C T33I2 T57M1 T28M1 T36C T51M1 T58M1 T45C T22M1 T50M1 T30M1 T32M1 T1M3 T15C T54M1 T10I1 T2C T15I1 T19C T24M1 T33M3 T16C T31M1 T11I1 T53M1 T37P1 T43I2 T4P1 T13I1 T47P2 T34I2 T2P1 T34M1 T52M1 T2M1 T55-56M1 T38M1 T5M1 T25M1 T26M1 0.10 0.20 0.30 0.40 BEca BEca 0.50 0.60 0.87 0.70 0.82 0.92 L’analyse des données 163 164 0.10 0.20 0.40 0.50 0.60 0.10 0.20 0.30 0.40 0.50 0.60 0.70 0.10 0.20 0.30 0.40 0.50 0.60 0.70 T30M1 T50M1 T22M1 T45C T58M1 T51M1 T36C T28M1 T57M1 T33I2 T18C T59M1 T29M1 T27M1 T23M1 T22P1 T20P1 T17I1 T17C T17P1 T8M1 T3C T2I1 T1I2 T32M1 T30M1 T50M1 T22M1 T45C T58M1 T51M1 T36C T28M1 T57M1 T33I2 T18C T59M1 T29M1 T27M1 T23M1 T22P1 T20P1 T17I1 T17C T17P1 T8M1 T3C T2I1 T1I2 T32M1 T30M1 T50M1 T22M1 T45C T58M1 T51M1 T36C T28M1 T57M1 T33I2 T18C T59M1 T29M1 T27M1 T23M1 T22P1 T20P1 T17I1 T17C T17P1 T8M1 T3C T2I1 T1I2 T32M1 T30M1 T50M1 T22M1 T45C T58M1 T51M1 T36C T28M1 T57M1 T33I2 T18C T59M1 T29M1 T27M1 T23M1 T22P1 T20P1 T17I1 T17C T17P1 T8M1 T3C T2I1 T1I2 0.00 T32M1 T1M3 T1M3 T1M3 0.00 T15C T1M3 T15C T15C T15C 0.00 T54M1 T54M1 T54M1 T54M1 0.00 T2C T10I1 T2C T10I1 T2C T10I1 T2C T10I1 T19C T15I1 T19C T15I1 T19C T24M1 T24M1 T24M1 T15I1 T24M1 T33M3 T16C T33M3 T16C T33M3 T19C T16C T33M3 T16C T15I1 T11I1 T31M1 T11I1 T31M1 T11I1 T31M1 T11I1 T31M1 T43I2 T43I2 T43I2 T37P1 T4P1 T4P1 T4P1 T53M1 T4P1 T43I2 T13I1 T13I1 T13I1 T37P1 T13I1 T47P2 T47P2 T47P2 T53M1 T47P2 T34I2 T34I2 T34I2 T37P1 T2P1 T34I2 T2P1 T2P1 T2P1 T53M1 T34M1 T34M1 T34M1 T34M1 T37P1 T2M1 T52M1 T2M1 T52M1 T2M1 T52M1 T2M1 T52M1 T53M1 T55-56M1 T55-56M1 0.70 T55-56M1 T55-56M1 0.30 T5M1 T38M1 T25M1 T38M1 0.76 T26M1 T38M1 T25M1 T38M1 T26M1 0.84 0.83 T5M1 T25M1 T26M1 BWun BWun T5M1 T25M1 MOC MOC T5M1 0.75 0.78 T26M1 BCca BCca 0.10 0.20 0.30 0.40 BRE BRE 0.50 0.60 0.70 Chapitre 7 0.60 0.70 T43I2 T43I2 0.50 T4P1 T4P1 T2P1 T27M1 T23M1 T22P1 T20P1 T17I1 T17C T17P1 T8M1 T3C T2I1 T1I2 T29M1 T27M1 T23M1 T22P1 T20P1 T17I1 T17C T17P1 T8M1 T3C T2I1 T1I2 0.00 T29M1 T59M1 0.00 T18C T59M1 T18C 0.00 T1I2 T2I1 T3C T8M1 T17P1 T17C T17I1 T20P1 T22P1 T23M1 T27M1 T29M1 T59M1 T18C T33I2 T57M1 T28M1 T33I2 T51M1 T45C T58M1 T57M1 T58M1 T58M1 T22M1 T33I2 T45C T45C T50M1 T36C T22M1 T22M1 T30M1 T28M1 T50M1 T50M1 T32M1 T57M1 T30M1 T30M1 T51M1 T32M1 T32M1 T15C T1M3 T36C T1M3 T1M3 T54M1 T51M1 T15C T15C T36C T54M1 T54M1 T10I1 T2C T15I1 T19C T24M1 T33M3 T11I1 T28M1 T2C T10I1 T15I1 T15I1 T2C T19C T19C T10I1 T24M1 T24M1 T16C T16C T33M3 T16C T33M3 T31M1 T11I1 T31M1 T11I1 T31M1 T37P1 T53M1 T37P1 T53M1 T37P1 T43I2 T4P1 T13I1 T47P2 T34I2 T53M1 0.40 T13I1 T13I1 0.30 T47P2 T47P2 0.20 T34I2 T34I2 0.10 T2P1 T2P1 0.70 T34M1 T34M1 0.60 T52M1 T52M1 0.50 T2M1 T2M1 0.40 T2M1 T52M1 T55-56M1 T55-56M1 0.30 T55-56M1 T38M1 T38M1 0.20 T5M1 T38M1 T5M1 0.10 T25M1 T25M1 T5M1 T34M1 T26M1 T26M1 T25M1 BCun BCun T26M1 BEun BEun 0.10 0.20 0.30 0.40 MaOr MaOr 0.50 0.60 0.70 L’analyse des données Figure 67 : Profil pour chaque groupe testé dans les analyses, élaboré à partir des expression count des dents clés retenues pour la Bohême (n=54). Liste des variables selon l’ordre dans le profil, de haut en bas: T26M1-T25M1-T5M1-T38M1-T55-56M1-T2M1-T52M1-T34M1-T2P1-T34I2-T47P2-T13I1-T4P1-T43I2-T37P1T53M1-T11I1-T31M1-T16C-T33M3-T24M1-T19C-T15I1-T2C-T10I1-T54M1-T15C-T1M3-T30M1-T32M1T50M1-T22M1-T45C-T36C-T51M1-T58M1-T28M1-T57M1-T33I2-T18C-T1I2-T2I1-T3C-T8M1-T17P1-T17CT17I1-T20P1-T22P1-T23M1-T27M1-T29M1-T59M1 165 Chapitre 7 7.2.1.1.4. Les analyses Les résultats des analyses multivariées sont des plus intéressants et montrent des tendances similaires (figure 68). -1 27 L’analyse ascendante hiérarchique complétée par un bootstrap présente deux regroupements à plus de 50% (figure 68a). 52 16 34 Le premier est composé par l’ensemble des groupes appartenant à la culture d’Unetice et par les Cordés orientaux (BEco). Au sein de cette configuration, un sous-groupe est formé par le Bronze ancien de Bohême centrale (BCun) et les Cordés orientaux. Le second est constitué des trois autres groupes cordés (VIK, BCco et BWco) et de l’ensemble des échantillons campaniformes. A l’intérieur, les Campaniformes des parties centrale (BCca) et occidentale (BWca) se mêlent aux ensembles cordés, tandis que le site de Mochov (MOC) se situant géographiquement dans la partie centrale de la Bohême forme un groupe fort et cohérent avec les Campaniformes orientaux (BEca). 84 100 91 8 57 32 80 34 32 ▼ 0.12 Le Bronze ancien présente à nouveau une concentration homogène. Les Cordés orientaux (BEco) sont également le groupe le plus proche de la culture d’Unetice. Les ensembles cordés et campaniformes, au contraire, montrent une certaine hétérogénéité. Un élément intéressant est la position du Cordé qui se place entre les Campaniformes et le Bronze ancien. L’analyse en composantes principales reflète à nouveau les éléments mis en évidence par les autres analyses. En effet, le nuage de points présente le regroupement des ensembles d’Unetice, la position plus éclatée du Campaniforme et du Cordé, ce dernier en intermédiaire entre les deux autres cultures (figure 68c). Le début du Bronze ancien, en revanche, révèle une forte cohésion des groupes étudiés indépendamment de leur localisation géographique, ce qui permet d’imaginer que l’on a affaire à un certain hermétisme aux influences externes. La position d’entre-deux du Cordé plaiderait en faveur de son implication tant au niveau du Campaniforme que de la culture d’Unetice dans leur mise en place. 166 -bBWca BEco ▼ BWco 0 BCun ■ ■ BRE MaOr ■ ■ ▼BCca ▼ BEun BEca ■ BWun VIK -0.3 -0.6 0 0.3 -c- VIK BWun 0.12 ■ BEun ■ ■ MaOr ■ BCun ■ BEca ▼ MOC ▼ BRE 0 BWco ▼BCca BEco BCco Que dire de ces résultats ? Les différentes analyses convergent et tendent à défendre l’idée de populations plus hétérogènes lors de l’Enéolithique final en Bohême qui traduirait probablement une mobilité ou des échanges populationnels plus accrus pendant ces périodes, qu’il faudrait, alors, préciser par les prochaines analyses (interrégionale, entre populations campaniformes ou sexes séparés). ■ BCun BEco ■ BWun ■ BEun ■ BRE ■ MaOr ▼ BEca ▼ MOC ▼ BWca BWco BCco VIK ▼ BCca BCco MOC Nous retrouvons les mêmes dispositions dans la représentation issue du multidimensional scaling complété par un minimal spanning tree (figure 68b). -a- 0 0 BWca ▼ -0.36 -0.3 0 Figure 68 : Résultats des multivariées pour la Bohême. 0.5 analyses a) Analyse ascendante hiérarchique, selon la méthode de Ward, complétée par un bootstrap . b) Multidimensional scaling complété par un minimal spanning tree (stress=0.09437). c) Analyse en composantes principales (axe 1=50.3% et axe 2=14.8%). L’analyse des données 7.2.1.2. La Hongrie L’intérêt de l’occupation campaniforme hongroise - le Campaniforme-Csepel - est multiple. Cette culture représente non seulement l’extension orientale du phénomène, mais elle est aussi, dans ce domaine, seule hors substrat cordé. Elle forme également une enclave dans la région de Budapest. 7.2.1.2.1. La constitution des groupes testés Le corpus se compose d’une centaine d’individus et de 15 sites localisés dans les parties centro-septentrionale et méridionale de la Hongrie (figure 69). Les ensembles appartiennent au Campaniforme-Csepel et aux cultures qui lui sont soit contemporaines (Obeda-Pitvaros, Nyirseg et Somogyvar-Vinkovci) soit successives (Nagyrev, Perjamos). Nous avons regroupé les sites par attribution culturelle pour avoir des effectifs intégrables dans les analyses. Deux groupes seront présentés lors de l’étape descriptive des données mais ne seront pas inclus dans les analyses. - les ensembles retenus pour les analyses (n suffisant) gisements - Campaniforme (19 individus) sites n° 1, 2, 3, 13 : CAM gisements - Nagyrev (35 individus) sites n° 4, 5, 10, 11 : NAG Figure 69 : La Hongrie, rappel de la localisation des sites (c.f. figure 10). gisements - Obedà-Pitvaros (10 individus) sites n° 6, 8, 12 : OB-PIT gisements - Perjamos (26 individus) sites n° 9, 14 : PER - les ensembles qui seront exclus des analyses (n insuffisant) gisement - Somogyvar-Vonkovci (2 individus) site n° 7 : S-V gisement - Nyirseg (2 individus) site n° 15 : NYI 7.2.1.2.2. Les incidences populationnelles Les incidences et les décomptes pour chaque variable se trouvent en table 33. Les données pour deux ensembles, Somogyvar-Vonkovci et Nyirseg, dont les effectifs sont nettement insuffisants (n=2), sont présentés à titre informatif. Les autres échantillons possèdent généralement suffisamment d’observations pour être intégrés dans les analyses. Lorsque les données sont insuffisantes pour un de ces groupes, nous éliminons la variable. Les traits observés sur l’ensemble des membres d’un district montrent, en général, des fréquences d’expression très proches les unes des autres. Un certain nombre de variables sont à expression constante. Il s’agit des traits rares appartenant au système FU-DTS et d’une partie des observations réalisées sur les racines. Les fréquences ajustées par la méthode de l’expression count E.C. pour chaque trait observé sur la dentition permanente sont données en table 34. 167 Chapitre 7 DENTS PERMANENTES Maxillaire 1 Absence congénitale 2 Nombre de radicules 3 Racine appendiciforme 4 Nombre de racines des prémolaires 5 Nombre de racines des molaires 6 Racine de Carabelli 7 Racine paramolaire 8 Racine idiopathique 9 Degré de rotation 10 Convexité vestibulaire 11 Sillon traversant 12 Tubercule distal 13 Cingulum vestibulaire 14 Gouttière vestibulaire 15 Crêtes linguales marginales 16 Tubercule dentaire 17 Crêtes vestibulaires marginales 18 Crête mésiale 19 Crête accessoire distale 20 Cuspides accessoires 21 Odontome 22 Extension de l'émail 23 Parastyle 24 Trait de Carabelli 25 Métacone 26 Hypocone 27 Métaconule 28 Tubercule distal accessoire 29 Tubercule mésial paracone 30 Tubercule mésial accessoire 31 Protoconule 32 Tubercule lingual paracone M3 M2 M1 P2 P1 C ● I2 I1 ● ● ● ● ● ● Mandibule ● ● ● ● ● ● ● ● ● ● ● ● ● 33 Absence congénitale 34 Nombre de radicules 35 Racine appendiciforme 36 Nombre racines canines 37 Racine de Tomes 38 Nombre racines molaires 39 Racine de Citroen 40 Racine paramolaire 41 Racine entomolaire 42 Racine idiopathique 43 Crêtes linguales marginales 44 Crêtes vestibulaires marginales 45 Crête accessoire distale 46 Odontome 47 Variation des cuspides linguales 48 Pli cuspide mésiolinguale 49 Fosse précuspidale 50 Crête trigonide intermédiaire 51 Crête trigonide distale 52 Nombre de cuspides 53 Dessin du sillon 54 Protostylide 55-56 Hypoconulide-Entoconulide 57 Métaconulide 58 Tubercule paracone 59 Tubercule de Citroen M3 M2 M1 P2 P1 C I2 I1 ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● Figure 70 : Représentation synthétique des dents clés retenues pour les analyses de la Hongrie (n=35). 7.2.1.2.3. Les variables mobilisées dans les analyses Pour la Hongrie, 35 variables sur les 79 traits localisés sur les dents clés ont été retenues. 20 d’entre elles s’observent sur le maxillaire et 15 sur la mandibule (figure 70). Les fréquences E.C. sont données table 35. Les traits localisés sur les couronnes ainsi que sur les canines et les molaires sont, en général, mieux représentés. Les profils d’expression des variables observées sur les dents clés mobilisables présentent des éléments intéressants (figure 71). Les profils appartenant aux cultures de Nagyrev et de Perjamos montrent une tendance assez proche. Il s’agit d’un élément intéressant puisque ces cultures sont toutes deux contemporaines. Le Campaniforme, quant à lui, possède un profil légèrement différent avec des expressions moins élevées des variables. La culture d’Obedà-Pitvaros, au contraire, possède des expressions sensiblement plus élevées pour certains traits. La culture de Nagyrev, même si elle est très proche de l’une des deux cultures méridionales hongroises, possède un profil qui la positionne en intermédiaire entre les différentes périodes étudiées. Profils d’expression des différents groupes définis pour la Hongrie Figure 71 : Profil pour chaque groupe testé dans les analyses, élaboré à partir des expression count des dents clés retenues pour la Hongrie (n=35). Liste des variables selon l’ordre dans le profil, de haut en bas: T26M1-T25M1-T5M1-T2M1-T55-56M1-T38M1-T52M1-T2P1-T34M1-T37P1-T4P1-T10I1-T28M1-T34P1-T47P2T16C-T24M1-T15C-T2C-T54M1-T43I2-T1M3-T33M3-T1I2-T14C-T17P1-T17C-T18C-T19C-T22M1-T34I2-T36CT45C-T46P2-T57M1 168 T47P2 T16C T24M1 T15C T2C T54M1 T47P2 T16C T24M1 T15C T2C T54M1 T47P2 T16C T24M1 T15C T2C T54M1 T36C T34I2 T22M1 T19C T18C T17C T17P1 T14C T1I2 T36C T34I2 T22M1 T19C T18C T17C T17P1 T14C T1I2 T36C T34I2 T22M1 T19C T18C T17C T17P1 T14C T1I2 0.00 T45C T45C T45C 0.00 T46P2 T46P2 T46P2 0.00 T1M3 T57M1 T1M3 T57M1 T1M3 T57M1 T43I2 T33M3 T43I2 T33M3 T43I2 T33M3 0.70 T34P1 T34P1 T34P1 0.60 T28M1 T28M1 T28M1 0.50 T10I1 T10I1 T10I1 0.40 T4P1 T4P1 T4P1 0.30 T37P1 T37P1 T37P1 0.20 T34M1 T34M1 T34M1 0.10 T2P1 T2P1 T2P1 0.70 T52M1 T52M1 T52M1 0.60 T38M1 T38M1 T38M1 0.50 T55-56M1 T55-56M1 T55-56M1 0.40 T2M1 T2M1 T2M1 0.30 T5M1 0.76 T5M1 T5M1 0.20 T25M1 0.79 T25M1 0.10 T26M1 0.80 T26M1 T25M1 NAG NAG T26M1 CAM CAM 0.10 0.20 0.30 OB-PIT 0.40 OB-PIT 0.50 0.60 0.70 0.00 T1I2 T14C T17P1 T17C T18C T19C T22M1 T34I2 T36C T45C T46P2 T57M1 T1M3 T33M3 T43I2 T54M1 T2C T15C T24M1 T16C T47P2 T34P1 T28M1 T10I1 T4P1 T37P1 T34M1 T2P1 T52M1 T38M1 T55-56M1 T2M1 T5M1 T25M1 0.97 1.00 T26M1 0.97 0.10 0.20 0.30 PER PER 0.40 0.50 0.60 0.70 0.73 0.80 0.80 L’analyse des données 169 Chapitre 7 7.2.1.2.4. Les analyses Les analyses se basent sur quatre ensembles seulement, ainsi la difficulté réside dans l’interprétation des configurations qu’elles proposent. Les résultats montrent cependant, en partie, des tendances similaires (figure 72). -1 ▼ L’analyse ascendante hiérarchique complétée par un bootstrap présente une situation intéressante (figure 72a). Nous avons un regroupement robuste (à 99%) des cultures contemporaines Nagyrev et Perjamos. La culture d’ObedàPitvaros, chronologiquement proche du Campaniforme, se rattache discrètement à ce regroupement. Le Campaniforme, en revanche, se détache complètement des autres périodes étudiées. CAM OB-PIT 100 73 ▼ 3 NAG 99 PER -b- Une configuration similaire, mais plus tranchée, est fournie par le nuage de points proposé par le multidimensional scaling complété par un minimal spanning tree (figure 72b). 0.24 Nous retrouvons la proximité des ensembles plus tardifs du Bronze ancien - Nagyrev et Perjamos - . Si le Campaniforme garde sa position éloignée, la culture d’Obedà-Pitvaros s’isole à son tour du regroupement principal. 0 L’analyse en composantes principales reflète les éléments mis en évidence par les deux premières analyses (figure 72c). La seule différence est que le regroupement principal n’est plus franchement observable. -a- 0 0 PER ▼ NAG ▼ CAM OB-PIT -0.24 -0.8 0 0.32 Que dire de ces résultats ? Le Nagyrev est une culture importante, tant dans l’impact ou l’influence au niveau matériel qu’il détient sur le Campaniforme, qu’au niveau de son extension géographique. Le Nagyrev semble, d’après les analyses, avoir une position forte et centrale des relations entre les différentes cultures étudiées. -c0.16 OB-PIT CAM ▼ NAG Le Campaniforme s'individualise sur l’ensemble des représentations. Il ne s’avère proche ni de la culture méridionale qui lui est contemporaine (Obedà-Pitvaros), ni du Nagyrev qui lui est en partie contemporain - dans sa phase ancienne - et qui lui succède. ▼ 0 PER -0.2 Nous pourrons éventuellement préciser la position éloignée du Campaniforme dans la prochaine analyse ou lors de l’étude des ensembles campaniformes. -0.48 0 Figure 72 : Résultats des multivariées pour la Hongrie. 0.64 analyses a) Analyse ascendante hiérarchique, selon la méthode de Ward, complétée par un bootstrap . b) Multidimensional scaling complété par un minimal spanning tree (stress=0.0). c) Analyse en composantes principales (axe 1=89.6% et axe 2=7.9%). 170 L’analyse des données 7.2.1.3. Les relations entre les ensembles du domaine oriental Une analyse des différents ensembles appartenant au domaine oriental est envisagée afin de comprendre les relations entre les différentes cultures des deux régions considérées : la Bohême et la Hongrie. 7.2.1.3.1. La constitution des groupes testés Nous avons inclus l’ensemble des groupes intégrés lors des analyses intrarégionales. La Bohême est constituée de 13 ensembles, pour rappel : - Cordé Bohême centrale (40 individus), sites n° 3, 16, 32, 36, 56, 59 : BCca Mochov (21 individus), site n° 25 : MOC Vikletice (90 individus), site n° 60 : VIK Bohême occidentale (58 individus), sites n° 1, 2, 8, 9, 10, 12, 14, 17, 19, 26, 27, 28, 40, 41, 42, 47, 49, 50, 51, 53, 55, 63 : BWco Bohême orientale (36 individus), sites n° 5, 7 : BEco Bohême centrale (24 individus), sites n° 4, 11, 15, 23, 29, 35, 37, 52, 54 : BCco - Unetice Bohême occidentale (38 individus), sites n° 2, 8, 22, 44, 45, 53, 58, 60, 61: BWun Brezno (38 individus), site n° 5 : BRE Bohême orientale (41 individus), sites n° 6, 13, 24, 39 : BEun Bohême centrale (36 individus), sites n° 23, 31, 33, 34, 35, 37, 38, 57 : BCun Mala Ohrada (47 individus), site n° 30 : MaOr - Campaniforme Bohême occidentale (20 individus), sites n° 18, 20, 42, 43, 44, 46, 48, 53, 62 : BWca Bohême orientale (17 individus), sites n° 5, 7, 21, 39 : BEca La Hongrie, quant à elle, est composée des 4 groupes intégrés dans l’analyse régionale, pour rappel : Gisements - Campaniforme (19 individus), sites n° 1, 2, 3, 13 : CAM Gisements - Nagyrev (35 individus), sites n° 4, 5, 10, 11 : NAG Gisements - Obedà-Pitvaros (10 individus), sites n° 6, 8, 12 : OB-PIT Gisements - Perjamos (26 individus), sites n° 9, 14 : PER DENTS PERMANENTES Maxillaire 1 Absence congénitale 2 Nombre de radicules 3 Racine appendiciforme 4 Nombre de racines des prémolaires 5 Nombre de racines des molaires 6 Racine de Carabelli 7 Racine paramolaire 8 Racine idiopathique 9 Degré de rotation 10 Convexité vestibulaire 11 Sillon traversant 12 Tubercule distal 13 Cingulum vestibulaire 14 Gouttière vestibulaire 15 Crêtes linguales marginales 16 Tubercule dentaire 17 Crêtes vestibulaires marginales 18 Crête mésiale 19 Crête accessoire distale 20 Cuspides accessoires 21 Odontome 22 Extension de l'émail 23 Parastyle 24 Trait de Carabelli 25 Métacone 26 Hypocone 27 Métaconule 28 Tubercule distal accessoire 29 Tubercule mésial paracone 30 Tubercule mésial accessoire 31 Protoconule 32 Tubercule lingual paracone M3 M2 M1 P2 P1 ● C ● ● I1 ● ● ● ● Mandibule ● ● ● ● ● ● ● ● ● ● ● I2 ● 33 Absence congénitale 34 Nombre de radicules 35 Racine appendiciforme 36 Nombre racines canines 37 Racine de Tomes 38 Nombre racines molaires 39 Racine de Citroen 40 Racine paramolaire 41 Racine entomolaire 42 Racine idiopathique 43 Crêtes linguales marginales 44 Crêtes vestibulaires marginales 45 Crête accessoire distale 46 Odontome 47 Variation des cuspides linguales 48 Pli cuspide mésiolinguale 49 Fosse précuspidale 50 Crête trigonide intermédiaire 51 Crête trigonide distale 52 Nombre de cuspides 53 Dessin du sillon 54 Protostylide 55-56 Hypoconulide-Entoconulide 57 Métaconulide 58 Tubercule paracone 59 Tubercule de Citroen M3 M2 M1 P2 P1 C I2 I1 ● ● ● ● ● ● ● ● ● ● ● Figure 73 : Représentation synthétique des dents clés retenues pour les analyses des ensembles appartenant au domaine oriental (n=30). 171 Chapitre 7 7.2.1.3.2. Les variables mobilisées dans les analyses Pour cette partie de l’étude, 30 variables sur les 79 traits localisés sur les dents clés ont été retenues. 19 d’entre elles s’observent sur le maxillaire et 11 sur la mandibule (figure 73). Les fréquences E.C. sont données table 36. Les traits localisés sur les couronnes et sur les molaires sont, en général, mieux représentés. 7.2.1.3.3. Les analyses Les analyses reprennent l’ensemble des groupes étudiés pour la Bohême et la Hongrie. Les résultats mêlent les configurations mises en évidence lors des études intrarégionales (figure 74). -1 -a- 0 0 L’analyse ascendante hiérarchique présente des regroupements qui ne sont pas vraiment robustes mais qui reflètent bien les résultats des analyses individuelles (figure 74a). Pour la Bohême, les deux regroupements - l’un réunissant les Unetices et les Cordés orientaux (BEco), l’autre mêlant les Cordés et les Campaniformes - se distinguent à nouveau. La position des groupes hongrois au sein de cette configuration est très intéressante puisque le Campaniforme se rattacherait modérément au premier groupe - Unetice et Cordé oriental - et les trois autres ensembles - Nagyrev, Obedà-Pitvaros et Perjamos - au second groupe constitué par le Campaniforme et le Cordé. Une configuration similaire est donnée par le nuage de points issu du multidimensional scaling (figure 74b). Pour la Bohême, nous retrouvons la proximité des ensembles du Bronze ancien, la position intermédiaire du Cordé et l’hétérogénéité du Campaniforme. Le Campaniforme hongrois se place à nouveau à proximité des ensembles unetices et les trois autres groupes s’unissent plutôt au Campaniforme tchèque ! L’analyse en composantes principales reproduit les éléments constatés lors des deux premières analyses (figure 74c). 12 11 21 25 2 21 22 8 11 100 18 14 16 24 33 58 83 10 OB-PIT BCco ▼ BCca VIK BWco ▼ NAG PER ▼ BEca ▼ MOC ■ BWun BEco ■ BCun ■ BRE ■ BEun ■ MaOr ▼ CAM ▼ BWca -b0.16 OB-PIT BWun ■ CAM ▼ ■ MaOr ■ ■ BEun ■ BCun BRE NAG ▼ VIK ▼BEca 0 BEco BWca▼ ▼BCca PER BWco MOC▼ BCco -0.2 -0.4 0 0.5 Que dire de ces résultats ? Nous aurions pu imaginer un regroupement des ensembles campaniformes hongrois et tchèques, mais cela n’a pas été le cas ! Si la situation semble évidente pour la Bohême, la Hongrie interroge. Figure 74 : Résultats des analyses multivariées pour le domaine oriental. a) Analyse ascendante hiérarchique, selon la méthode de Ward, complétée par un bootstrap . b) Multidimensional scaling complété par un minimal spanning tree (stress=0.068). c) Analyse en composantes principales (axe 1=73.3% et axe 2=10.3%). 172 -cBCco 0.25 BEca▼ BWco MOC▼ BCca▼ PER 0 ▼ NAG ▼ BWca BEco BCun ■ CAM BRE ■ ■ BEun▼ ■ VIK MaOr OB-PIT BWun ■ -0.24 -0.64 0 0.32 L’analyse des données 7.2.2. Le domaine méridional L’aire méridionale est un territoire où les modalités de l’émergence du Campaniforme sera vraisemblablement plus délicate à saisir. En effet, la spécificité des vestiges campaniformes et leur complexité relève d’un exercice des plus laborieux. 7.2.2.1. Le sud de la France Les données archéologiques révèlent généralement deux phases d’occupation campaniforme. La première marque une certaine rupture avec les traditions locales, la seconde est une période d’intégration et de cohabitation. Les contextes funéraires complexes, d’un point de vue stratigraphique, dans cette région n’ont pas induit une multitude d’études anthropologiques dans une perspective d’histoire du peuplement. 7.2.2.1.1. La constitution des groupes testés L’échantillon comporte près de 700 individus et 5 sites localisés en grande partie dans la partie orientale de la région (figure 75). Les gisements se situent entre le Néolithique final/Chalcolithique et le Bronze ancien. Nous avons uniquement affaire à des sépultures collectives. Les attributions culturelles ne sont pas toujours claires, ainsi nous n’avons pas toujours pu identifier et travailler avec des groupes culturels ou périodes distinctes. - les ensembles sélectionnés Boileau - Néolithique final (130 individus), site n° 4 : BOI Peyraoutes - Chalcolithique/Campaniforme (172 individus), site n° 2 : PEY Roaix, couche 5 - Chalcolithique (92 individus), site n° 3 : ROc5 Roaix, couche 2 - Chalcolithique (49 individus), site n° 3 : ROc2 Figure 75 : Le sud de la France, rappel de la localisation des sites (c.f. figure 11). Villedubert - Néolithique final (18 individus), site n° 1 : VILn Villedubert - Campaniforme (70 individus), site n° 1 : VILca Villedubert - phase indéterminée (19 individus), site n° 1 : VILind Cudières - Chalcolithique/Bronze ancien (98 individus), site n° 5 : CUD 7.2.2.1.2. Les incidences populationnelles Les incidences et les décomptes pour chaque variable se trouvent en table 37 pour la dentition permanente et en table 38 pour la dentition déciduale. Seul l’échantillon attribué au Néolithique final du site de Villedubert (n°1 - VILn) présente un nombre insuffisant d’observations, nous décidons ainsi de ne pas l’inclure au sein des analyses. Les autres groupes possèdent, pour la plupart, assez d’observations pour être intégrés dans les analyses. Lorsque les données sont insuffisantes pour l’un de ces ensembles, nous éliminons de préférence la variable. Pour la dentition permanente (table 37), les traits observés sur l’ensemble des membres d’un district présentent des fréquences d’expression comparables. Les variables appartenant au système FU-DTS et les traits secondaires observés sur les racines ne montrent que très peu de variations. Nous avons gardé pour la dentition déciduale les groupes définis pour la dentition permanente (table 38). Les effectifs sont faibles et la majorité des variables est constante. Les traits mobilisables ne révèlent pas de grandes différences entre les groupes étudiés. 173 Chapitre 7 Les fréquences ajustées par la méthode de l’expression count E.C. pour chaque trait observé sur la dentition permanente sont données en table 39. DENTS PERMANENTES Maxillaire 1 Absence congénitale 2 Nombre de radicules 3 Racine appendiciforme 4 Nombre de racines des prémolaires 5 Nombre de racines des molaires 6 Racine de Carabelli 7 Racine paramolaire 8 Racine idiopathique 9 Degré de rotation 10 Convexité vestibulaire 11 Sillon traversant 12 Tubercule distal 13 Cingulum vestibulaire 14 Gouttière vestibulaire 15 Crêtes linguales marginales 16 Tubercule dentaire 17 Crêtes vestibulaires marginales 18 Crête mésiale 19 Crête accessoire distale 20 Cuspides accessoires 21 Odontome 22 Extension de l'émail 23 Parastyle 24 Trait de Carabelli 25 Métacone 26 Hypocone 27 Métaconule 28 Tubercule distal accessoire 29 Tubercule mésial paracone 30 Tubercule mésial accessoire 31 Protoconule 32 Tubercule lingual paracone M3 M2 M1 P2 P1 ● C ● ● I2 I1 ● ● ● ● ● Mandibule ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● 33 Absence congénitale 34 Nombre de radicules 35 Racine appendiciforme 36 Nombre racines canines 37 Racine de Tomes 38 Nombre racines molaires 39 Racine de Citroen 40 Racine paramolaire 41 Racine entomolaire 42 Racine idiopathique 43 Crêtes linguales marginales 44 Crêtes vestibulaires marginales 45 Crête accessoire distale 46 Odontome 47 Variation des cuspides linguales 48 Pli cuspide mésiolinguale 49 Fosse précuspidale 50 Crête trigonide intermédiaire 51 Crête trigonide distale 52 Nombre de cuspides 53 Dessin du sillon 54 Protostylide 55-56 Hypoconulide-Entoconulide 57 Métaconulide 58 Tubercule paracone 59 Tubercule de Citroen M3 M2 M1 P2 P1 C I2 I1 ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● Figure 76 : Représentation synthétique des dents clés retenues pour les analyses du sud de la France (n=57). 7.2.2.1.3. Les variables mobilisées dans les analyses Pour le sud de la France, 57 variables sur les 79 traits localisés sur les dents clés ont été retenues. 35 d’entre elles s’observent sur le maxillaire et 22 sur la mandibule (figure 76). Les fréquences E.C. qui seront utilisées dans les analyses sont données en table 40. Les traits localisés sur les couronnes sont, en général, mieux représentés ainsi que les variables observées sur les molaires. Les profils d’expression des variables observées sur les dents clés mobilisables permettent d’isoler des spécificités selon les groupes (figure 77). Les profils du site des Peyraoutes (n°2 - PEY) et de Cudières (n°5 - CUD) sont assez proches et se différencient par une fréquence peu élevée T5, T25 et T26 par rapport aux autres ensembles. Les profils appartenant à des périodes ou couches distinctes dans un même gisement, tels Villedubert (n°1 - VIL) ou Roaix (n°3 - RO), sont intéressants. On retrouve, dans les grandes lignes, les mêmes tendances avec quelques particularités de fréquences d’expression. Pour Roaix, on distingue pour la couche 2 (ROc2) par rapport à la couche 5 (ROc5) des fréquences élevées de T2 et bien inférieures pour T13, T16 et T55-56. De même, pour Villedubert, l’occupation campaniforme (VILca) présente des taux plus élevés de T2, T5 et T53. Le gisement des Boileau (n°4 - BOI) semble très proche de cette séquence campaniforme. Profils d’expression des différents groupes définis pour le sud de la France 174 0.20 0.30 0.40 0.50 0.60 0.70 0.10 0.20 0.30 0.50 0.60 0.70 0.10 0.20 0.30 T2P1 T16C T16C T15I1 T11I1 T11I1 0.50 0.60 0.70 T57M1 T46P2 T57M1 T46P2 T57M1 T46P2 T21P1 T21P1 T17P1 T35C T31M1 T27M1 T23M1 T22P1 T21P1 T17P1 T14I1 T7M1 T3I1 T1P2 T59M1 T51M1 T35C T31M1 T27M1 T23M1 T22P1 T21P1 T17P1 T14I1 T7M1 T3I1 T1P2 0.00 T22P1 T22P1 T17I1 T17I1 0.00 T51M1 T27M1 T23M1 T27M1 T23M1 0.00 0.00 T3I1 T1P2 T3I1 T7M1 T1P2 T14I1 T7M1 T14I1 T17P1 T31M1 T31M1 T35C T59M1 T51M1 T51M1 T35C T59M1 T59M1 T18C T17I1 T17I1 T18C T45C T18C T18C T45C T45C T45C T17C T43I1 T43I1 T17C T43I1 T17C T43I1 T17C T58M1 T46P2 T58M1 T57M1 T19C T50M1 T19C T58M1 T19C T50M1 T19C T50M1 T32M1 T32M1 T58M1 T50M1 T20P1 T20P1 T20P1 T22M1 T22M1 T22M1 T32M1 T16I1 T22M1 T16I1 T16I1 T20P1 T28M1 T28M1 T28M1 T16I1 T32M1 T30M1 T30M1 T30M1 T28M1 T36C T30M1 T36C T1I2 T36C T1I2 T11I1 T15I1 T16C T54M1 T1I2 T1I2 T15C T34P1 T36C T11I1 T15I1 T54M1 T54M1 T16C T54M1 T34P1 T15C T13I1 T37P1 T29M1 T13I1 T29M1 T15I1 T15C T34P1 T15C T34P1 T13I1 T29M1 T13I1 T29M1 T24M1 T37P1 T37P1 T37P1 T24M1 T24M1 T24M1 T2C T53M1 T53M1 T2C T53M1 T2C T2P1 T34I2 T33M3 T2C T34I2 T33M3 T1M3 T10I1 T1M3 T47P2 T55-56M1 T34M1 T52M1 T10I1 0.40 T34C T2M1 T47P2 T55-56M1 T34M1 T52M1 T2M1 T53M1 0.40 T1M3 T1M3 0.10 T10I1 T10I1 T2P1 T47P2 T47P2 T34I2 T55-56M1 T55-56M1 T33M3 T34M1 T34M1 T2P1 T52M1 T52M1 T34I2 T2M1 T2M1 T33M3 T5M1 T38M1 T34C T34C T34C T38M1 T38M1 T38M1 T25M1 T5M1 T5M1 T5M1 T26M1 T25M1 T25M1 T26M1 T26M1 ROc5 ROc5 T25M1 PEY PEY T26M1 BOI BOI 0.10 0.20 0.30 0.40 ROc2 ROc2 0.50 0.60 0.70 L’analyse des données 175 176 0.10 0.20 0.30 0.40 0.50 0.60 0.70 0.10 0.20 0.30 0.50 0.60 0.70 T47P2 T10I1 T1M3 T2P1 T34I2 T2C T53M1 T24M1 T37P1 T13I1 T29M1 T15C T47P2 T10I1 T1M3 T2P1 T33M3 T34I2 T2C T53M1 T24M1 T37P1 T13I1 T29M1 T47P2 T10I1 T1M3 T2P1 T33M3 T34I2 T2C T53M1 T24M1 T37P1 T13I1 T29M1 0.40 T34M1 T34M1 T1I2 T36C T30M1 T16I1 T22M1 T20P1 T32M1 T50M1 T19C T58M1 T57M1 T46P2 T17C T43I1 T1I2 T36C T30M1 T28M1 T16I1 T22M1 T20P1 T32M1 T50M1 T19C T58M1 T57M1 T46P2 T17C T43I1 T30M1 T28M1 T16I1 T22M1 T20P1 T32M1 T50M1 T19C T58M1 T57M1 T46P2 T17C T43I1 T45C Figure 77 : Profil pour chaque groupe testé dans les analyses, élaboré à partir des expression count des dents clés retenues pour le sud de la France (n=57). Liste des variables selon l’ordre dans le profil, de haut en bas: T26M1-T25M1-T5M1-T38M1-T34C-T2M1-T52M1-T34M1-T55-56M1-T47P2-T10I1-T1M3-T2P1-T33M3-T34I2T2C-T53M1-T24M1-T37P1-T13I1-T29M1-T15C-T34P1-T54M1-T16C-T15I1-T11I1-T1I2-T36C-T30M1-T28M1T16I1-T22M1-T20P1-T32M1-T50M1-T19C-T57M1-T58M1-T46P2-T17C-T43I1-T45C-T18C-T17I1-T1P2-T3I1T7M1-T14I1-T17P1-T21P1-T22P1-T23M1-T27M1-T31M1-T35C-T51M1-T59M1 T35C T31M1 T27M1 T23M1 T22P1 T21P1 T17P1 T14I1 T7M1 T3I1 T1P2 T51M1 T35C T31M1 T27M1 T23M1 T22P1 T21P1 T17P1 T14I1 T7M1 T3I1 T1P2 T51M1 T35C T31M1 T27M1 T23M1 T22P1 T21P1 T17P1 T14I1 T7M1 T3I1 T1P2 0.00 T51M1 T59M1 T59M1 0.00 T18C T59M1 T18C 0.00 T45C T17I1 T45C T17I1 T18C T17I1 T28M1 T11I1 T11I1 T1I2 T11I1 T36C T16C T15I1 T16C T15I1 T16C T15I1 T34P1 T54M1 T15C T34P1 T54M1 T15C T34P1 T54M1 T33M3 T52M1 T55-56M1 T52M1 T55-56M1 T34M1 T52M1 T34C T2M1 T55-56M1 T34C T38M1 T38M1 T2M1 T5M1 T38M1 T5M1 T34C T25M1 T25M1 T5M1 T2M1 T26M1 T26M1 T25M1 VILind VILind T26M1 VILca VILca 0.10 0.20 0.30 0.40 CUD CUD 0.50 0.60 0.70 Chapitre 7 L’analyse des données 7.2.2.1.4. Les analyses Les résultats des analyses multivariées proposent des configurations proches (figure 78). -1 L’analyse ascendante hiérarchique complétée par un bootstrap présente deux regroupements robustes (figure 78a). 99 Le second est constitué des trois autres gisements, toutes phases d’occupation confondues. A l’intérieur de ce regroupement, la première phase du site de Roaix (ROc5) s’isole des autres. Le site des Boileau (BOI) se rattache à l’occupation campaniforme de Villedubert (VILca), tandis que la seconde couche de Roaix (ROc2) s’approche de l’ensemble indéterminé de Villedubert (VILind). Il faut mentionner l’hétérogénéité des phases du Néolithique final ou du Chalcolithique avec les sites de Roaix et des Boileau. L’analyse en composantes principales reflète à nouveau les éléments mis en évidence par les autres analyses. En effet, le nuage de points présente le regroupement des ensembles récents et la variabilité des groupes antérieurs (figure 78c). CUD ROc5 4 72 71 BOI ▼ VILca 51 VILind 44 ROc2 -b0.24 ROc5 Les mêmes tendances se dessinent dans la représentation issue du multidimensional scaling complété par un minimal spanning tree (figure 78b). La proximité des sites récents - Les Peyraoutes (PEY) et Cudières (CUD) - est à nouveau identifiable. Ce regroupement se rattache à l’occupation campaniforme de Villedubert (VILca). Celle-ci est également reliée à d’autres ensembles, comme la phase indéterminée du même site (VILind) ou le site des Boileau (BOI). ▼ PEY ■ 100 Le premier est constitué par deux ensembles tardifs : Les Peyraoutes (PEY) - Chalcolithique/Campaniforme - et Cudières (CUD) - Chalcolithique/Bronze ancien -. -a- 0 0 CUD VILca VILind 0 ▼ ■ ▼ PEY BOI ROc2 -0.3 -0.48 0 0.48 -c ROc2 0.32 VILind Que dire de ces résultats ? Les gisements du Néolithique final et du Chalcolithique présentent, à l’évidence, une situation plus hétéroclite. Le site de Roaix en est un bon exemple, dans la mesure où ses deux occupations (couches 5 et 2) ne semblent pas refléter un seul grand ensemble homogène, mais plutôt deux populations distinctes. En revanche, les gisements récents de Les Peyraoutes et de Cudières, contenant un mélange d’occupation chalcolithique avec une phase campaniforme pour le premier et bronze ancien pour le second, s’isolent en gardant une proximité avec l’ensemble campaniforme de Villedubert. Ce dernier site, pour lequel nous avons analysé un groupe campaniforme et un ensemble indéterminé, semble se positionner entre ces deux courants. BOI 0 CUD PEY▼ ■ VILca ▼ -0.24 ROc5 -0.48 0 0.64 Figure 78 : Résultats des analyses multivariées pour le sud de la France. a) Analyse ascendante hiérarchique, selon la méthode de Ward, complétée par un bootstrap . b) Multidimensional scaling complété par un minimal spanning tree (stress=0.08861). c) Analyse en composantes principales (axe 1=68.3% et axe 2=13.7%). 177 Chapitre 7 7.2.2.2. Le nord de l’Espagne La péninsule Ibérique est une région importante dans la compréhension du phénomène Campaniforme. Néanmoins, le contexte funéraire difficile ne nous permet pas une analyse très fine. 7.2.2.2.1. La constitution des groupes testés L’échantillon est constitué de près de 180 individus et 14 sites situés dans la partie septentrionale de la Meseta (figure 79). Les ensembles étudiés sont à rattacher au Néolithique final, au Chalcolithique, au Campaniforme et à l’âge du Bronze. Nous avons décidé de garder les gisements comportant plus de 18 individus et de regrouper les autres sites selon leur attribution culturelle. Nous avons 3 ensembles pour le Néolithique final, 2 pour le Chalcolithique, 1 pour le Campaniforme et 1 pour l’âge du Bronze. Un gisement présente un mélange non distinguable appartenant au Néolithique final et au Campaniforme. - Ensembles inclus dans les analyses (n suffisant) La Sima - Néolithique final (22 individus) site n° 8 : LSnf La Velilla - Néolithique final (50 individus) site n° 9 : LVnf Figure 79 : Le nord de l’Espagne, rappel de la localisation des sites (c.f. figure 12). Los Zumacalles - Néolithique final (18 ind.) site n° 12 : LZnf Valle de las Higueras - Chalcolithique (38 individus), site n° 13 : VHcha Las Arnillas - Néolithique final/Campaniforme (24 individus), site n°10 : LAnf/ca Gisements - Campaniforme (18 individus) sites n° 3, 7, 8, 13 : DIVca Gisements - Chalcolithique (13 individus), sites n° 2, 6, 11, 14 : DIVcha - Ensembles exclus des analyses (n insuffisant) Gisements - Bronze (5 individus) sites n° 1, 4, 5 : DIVbz 7.2.2.2.2. Les incidences populationnelles Les incidences et les décomptes pour chaque variable se trouvent en table 41. L’échantillon constitué des individus de l’âge du Bronze (DIVbz) ne contient pas suffisamment d’observations, nous décidons ainsi de ne pas l’inclure au sein des analyses multivariées. Les autres groupes possèdent, pour la plupart, assez d’observations pour être intégrés dans les analyses. Lorsque les données sont insuffisantes pour l’un de ces ensembles, nous éliminons de préférence la variable. Pour la dentition permanente, les traits observés sur l’ensemble des membres d’un district présentent, en général, des fréquences d’expression similaires (table 41). Sans grand étonnement, les traits s’exprimant sur les racines et appartenant au système FU-DTS (traits rares) révèlent une importante constance d’expression. Les fréquences ajustées par la méthode de l’expression count E.C. pour chaque trait observé sur la dentition permanente sont données en table 42. 178 L’analyse des données DENTS PERMANENTES Maxillaire 1 Absence congénitale 2 Nombre de radicules 3 Racine appendiciforme 4 Nombre de racines des prémolaires 5 Nombre de racines des molaires 6 Racine de Carabelli 7 Racine paramolaire 8 Racine idiopathique 9 Degré de rotation 10 Convexité vestibulaire 11 Sillon traversant 12 Tubercule distal 13 Cingulum vestibulaire 14 Gouttière vestibulaire 15 Crêtes linguales marginales 16 Tubercule dentaire 17 Crêtes vestibulaires marginales 18 Crête mésiale 19 Crête accessoire distale 20 Cuspides accessoires 21 Odontome 22 Extension de l'émail 23 Parastyle 24 Trait de Carabelli 25 Métacone 26 Hypocone 27 Métaconule 28 Tubercule distal accessoire 29 Tubercule mésial paracone 30 Tubercule mésial accessoire 31 Protoconule 32 Tubercule lingual paracone M3 M2 M1 P2 P1 C I2 I1 ● ● ● ● ● Mandibule ● ● ● ● ● ● 33 Absence congénitale 34 Nombre de radicules 35 Racine appendiciforme 36 Nombre racines canines 37 Racine de Tomes 38 Nombre racines molaires 39 Racine de Citroen 40 Racine paramolaire 41 Racine entomolaire 42 Racine idiopathique 43 Crêtes linguales marginales 44 Crêtes vestibulaires marginales 45 Crête accessoire distale 46 Odontome 47 Variation des cuspides linguales 48 Pli cuspide mésiolinguale 49 Fosse précuspidale 50 Crête trigonide intermédiaire 51 Crête trigonide distale 52 Nombre de cuspides 53 Dessin du sillon 54 Protostylide 55-56 Hypoconulide-Entoconulide 57 Métaconulide 58 Tubercule paracone 59 Tubercule de Citroen M3 M2 M1 P2 P1 C I2 I1 ● ● ● ● ● ● Figure 80 : Représentation synthétique des dents clés retenues pour les analyses du nord de l’Espagne (n=17). 7.2.2.2.3. Les variables mobilisées dans les analyses Pour le nord de l’Espagne, seules 17 variables sur les 79 traits localisés sur les dents clés ont été retenues. 11 d’entre elles s’observent sur le maxillaire et 6 sur la mandibule (figure 80). Les fréquences E.C. sont données table 43. Les traits localisés sur les couronnes ainsi que sur les incisives, les canines et les molaires sont, en général, mieux représentés. Les variables mobilisables ne sont pas nombreuses (n=17) néanmoins les profils d’expression révèlent des éléments intéressants. Des divergences entre les ensembles peuvent être mises en évidence (figure 81). La Sima (LSnf) et La Vellila (LVnf) sont très proches, ainsi que l’échantillon campaniforme (DIVca) et l’occupation chalcolithique de la Valle de las Higueras (VHcha). Los Zumacalles se distingue de ces ensembles par la fréquence plus élevée de trois traits (nombre de racines des molaires supérieures - T5 -, métacone - T25 - et hypocone - T26 -). Les profils des deux derniers ensembles (LAnf/ca et DIVcha) diffèrent sensiblement des autres groupes. Ils possèdent des fréquences élevées pour quatre variables (hypoconulide-entoconulide - T55-56 -, nombre de cuspides - T52 -, nombre de racines des molaires inférieures - T38 - et variations des cuspides linguales - T47 -). Las Arnillas (LAnf/ca), qui est un site où les différentes occupations néolithique final et campaniforme ne se distinguent pas, possède également des fréquences élevées pour le nombre de radicules de la première molaire inférieure - T34 -. Le groupe formé des divers sites chalcolithiques (DIVcha) présente des incidences élevées pour deux traits supplémentaires. Il s’agit de l’absence congénitale de l’incisive latérale supérieure (T2) et le nombre de radicules de la première prémolaire supérieure (T2). 179 180 0.60 0.70 0.10 0.20 0.40 0.50 0.60 0.70 0.20 Figure 81 : Profil de chaque groupe testé dans les analyses, élaboré à partir des expression count des dents clés retenues pour le nord de l’Espagne (n=17). Liste des variables selon l’ordre dans le profil, de haut en bas: T26M1-T25M1-T5M1-T55-56M1-T52M1-T38M1-T34M1-T2P2-T2P1-T47P2-T2C-T16C-T54M1-T28M1-T1I2T3C-T17P1-T17C 0.50 0.60 0.70 T17P1 T3C T1I2 T17P1 T3C T1I2 0.40 T17C T17C 0.00 T3C T28M1 T28M1 0.00 T17P1 T54M1 T54M1 T2C 0.00 T1I2 T17C T28M1 T54M1 T16C T2C T16C T2C T47P2 T2P2 T2P1 T34M1 T38M1 T52M1 T55-56M1 T16C 0.30 T47P2 T47P2 0.10 T2P2 T2P2 0.70 T2P1 T2P1 0.60 T34M1 T34M1 0.50 T38M1 T38M1 0.40 T52M1 T52M1 0.30 T55-56M1 T55-56M1 0.20 T5M1 T5M1 0.10 T25M1 T25M1 T5M1 T26M1 T26M1 VHcha VHcha 0.00 T25M1 DIVcha DIVcha 0.00 T26M1 0.00 0.10 0.20 0.30 0.40 0.50 0.60 0.70 0.10 0.20 0.30 DIVca 0.40 DIVca 0.50 0.60 0.70 0.00 T3C T1I2 T3C T1I2 T17P1 T17P1 T17P1 T3C T17C T17P1 T17C T17C T17C T1I2 T28M1 T28M1 T28M1 T28M1 T3C T54M1 T54M1 T54M1 T54M1 T1I2 T2C T16C T2C T16C T2C T16C T2C T16C 0.30 T47P2 T47P2 T47P2 0.50 T2P2 T47P2 T2P2 T2P2 T2P2 0.40 T2P1 T2P1 T2P1 T2P1 0.30 T34M1 T34M1 T34M1 T34M1 0.20 T38M1 T38M1 T38M1 T38M1 0.10 T52M1 T52M1 T52M1 T52M1 T5M1 T55-56M1 T55-56M1 T55-56M1 T55-56M1 T25M1 T5M1 T5M1 T25M1 T25M1 T26M1 T5M1 LZnf LZnf T26M1 T25M1 LVnf LVnf T26M1 LSnf LSnf T26M1 0.10 0.20 0.30 LAnf/ca 0.40 LAnf/ca 0.50 0.60 0.70 Chapitre 7 Profils d’expression des différents groupes définis pour le nord de l’Espagne L’analyse des données 7.2.2.2.4. Les analyses Les résultats des analyses multivariées sont des plus intéressants (figure 82). -1 L’analyse ascendante hiérarchique complétée par un bootstrap présente deux regroupements robustes (figure 82a). Le premier est composé par les échantillons datant du Néolithique final (LZnf, LSnf, LVnf), par les Campaniformes (DIVca) et le Chalcolithique de la Valle de Las Higueras (VHcha). Au sein de cette configuration, les ensembles récents se regroupent, ainsi que deux des trois groupes du Néolithique final (LSnf et LVnf). Le second est constitué des deux derniers échantillons, soit par la sépulture collective de Las Arnillas mêlant Néolithique final et Campaniforme (LAnf/ca) et par un ensemble chalcolithique (DIVcha). Nous retrouvons les mêmes dispositions dans la représentation issue du multidimensional scaling complété par un minimal spanning tree (figure 82b). Les sites datant du Néolithique final ne sont pas proches sur la configuration, ils sont néanmoins reliés entre eux. Le gisement de la Velilla (LVnf) se situe en position intermédiaire, avec de part et d’autre, les échantillons récents (LAnf/ca, DIVca, DIVcha et VHcha). A noter : les ensembles campaniformes (LAnf/ca et DIVca) se placent entre le Néolithique final et le Chalcolithique. L’analyse en composantes principales reflète à nouveau les éléments mis en évidence par les autres analyses. En effet, le nuage de points présente le regroupement des ensembles du Néolithique final (LZnf, LSnf, LVnf) et l’hétérogénéité des échantillons postérieurs (LAnf/ca, DIVca, DIVcha et VHcha) (figure 82c). ▼ DIVcha 100 LZnf 4 86 ▼ VHcha 63 ▼ DIVca 62 LSnf 37 LVnf -b- LAnf/ca ▼ 0.24 LVnf ▼ DIVca ▼ LSnf 0 VHcha ▼ DIVcha LZnf -0.3 -0.48 0 0.48 -c▼ LAnf/ca 0.32 LVnf 0 ▼ ▼ DIVca VHcha LSnf DIVcha ▼ Les différentes analyses dépeignent une situation intéressante de la fin du Néolithique de la Meseta nord. -0.32 Les ensembles récents, en revanche, semblent montrer une plus grande variabilité. De plus, un élément intriguant est la position des Campaniformes en intermédiaire entre les populations du Néolithique final et du Chalcolithique. ▼ LAnf/ca 73 Que dire de ces résultats ? Les différents groupes appartenant au Néolithique final paraissent homogènes entre eux. -a- 0 0 -0.48 LZnf 0 0.6 Figure 82 : Résultats des analyses multivariées pour le nord de l’Espagne. a) Analyse ascendante hiérarchique, selon la méthode de Ward, complétée par un bootstrap . b) Multidimensional scaling complété par un minimal spanning tree (stress=0.0). c) Analyse en composantes principales (axe 1=68.4% et axe 2=25.1%). 181 Chapitre 7 7.2.2.3. Les relations entre les ensembles du domaine méridional Une analyse des différents ensembles appartenant au domaine méridional est envisagée afin de comprendre les relations entre les différentes cultures des deux régions considérées : le sud de la France et le nord de l’Espagne. 7.2.2.3.1. La constitution des groupes testés Nous avons inclus l’ensemble des groupes intégrés lors des analyses intrarégionales, à l’exception du site de La Sima (LSnf) pour le nord de l’Espagne pour lequel nous n’avons pas un nombre suffisant d’observations. Le sud de la France est constitué de 7 ensembles, pour rappel : Boileau - Néolithique final (130 individus), site n° 4 : BOI Villedubert - Campaniforme (70 individus), site n° 1 : VILca Peyraoutes - Chalcolithique/Campaniforme (172 individus), site n° 2 : PEY Villedubert - phase indéterminée (19 individus), site n° 1 : VILind Roaix, couche 5 - Chalcolithique (92 individus), site n° 3 : ROc5 Cudières - Chalcolithique/Bronze ancien (98 individus), site n° 5 : CUD Roaix, couche 2 - Chalcolithique (49 individus), site n° 3 : ROc2 Le nord de l’Espagne est composé de 6 groupes, pour rappel : La Velilla - Néolithique final (50 individus), site n° 9 : LVnf Gisements - Chalcolithique (13 individus), sites n° 2, 6, 11, 14 : DIVcha Los Zumacalles - Néolithique final (18 individus), site n° 12 : LZnf Valle de las Higueras - Chalcolithique (38 individus), site n° 13 : VHcha Las Arnillas - Néolithique final/Campaniforme (24 individus), site n°10 : LAnf/ca Gisements - Campaniforme (18 individus), sites n° 3, 7, 8, 13 : DIVca DENTS PERMANENTES Maxillaire 1 Absence congénitale 2 Nombre de radicules 3 Racine appendiciforme 4 Nombre de racines des prémolaires 5 Nombre de racines des molaires 6 Racine de Carabelli 7 Racine paramolaire 8 Racine idiopathique 9 Degré de rotation 10 Convexité vestibulaire 11 Sillon traversant 12 Tubercule distal 13 Cingulum vestibulaire 14 Gouttière vestibulaire 15 Crêtes linguales marginales 16 Tubercule dentaire 17 Crêtes vestibulaires marginales 18 Crête mésiale 19 Crête accessoire distale 20 Cuspides accessoires 21 Odontome 22 Extension de l'émail 23 Parastyle 24 Trait de Carabelli 25 Métacone 26 Hypocone 27 Métaconule 28 Tubercule distal accessoire 29 Tubercule mésial paracone 30 Tubercule mésial accessoire 31 Protoconule 32 Tubercule lingual paracone 182 M3 M2 M1 P2 P1 ● C ● ● I1 ● ● ● ● Mandibule ● ● ● ● ● ● ● ● ● ● ● I2 ● 33 Absence congénitale 34 Nombre de radicules 35 Racine appendiciforme 36 Nombre racines canines 37 Racine de Tomes 38 Nombre racines molaires 39 Racine de Citroen 40 Racine paramolaire 41 Racine entomolaire 42 Racine idiopathique 43 Crêtes linguales marginales 44 Crêtes vestibulaires marginales 45 Crête accessoire distale 46 Odontome 47 Variation des cuspides linguales 48 Pli cuspide mésiolinguale 49 Fosse précuspidale 50 Crête trigonide intermédiaire 51 Crête trigonide distale 52 Nombre de cuspides 53 Dessin du sillon 54 Protostylide 55-56 Hypoconulide-Entoconulide 57 Métaconulide 58 Tubercule paracone 59 Tubercule de Citroen M3 M2 M1 P2 P1 C I2 I1 ● ● ● ● ● ● ● ● ● ● ● Figure 83 : Représentation synthétique des dents clés retenues pour les analyses des ensembles appartenant au domaine méridional (n=30). L’analyse des données 7.2.2.3.2. Les variables mobilisées dans les analyses Pour cette partie de l’étude, 30 variables sur les 79 traits localisés sur les dents clés ont été retenues. 19 d’entre elles s’observent sur le maxillaire et 11 sur la mandibule (figure 83). Les fréquences E.C. sont données table 44. Les traits localisés sur les couronnes des molaires sont en général mieux représentés. 7.2.2.3.3. Les analyses Les analyses reprennent une large majorité des groupes étudiés pour le sud de la France et le nord de l’Espagne. Les résultats mélangent les configurations mises en évidence lors des études individuelles (figure 84). -1 L’analyse ascendante hiérarchique complétée par un bootstrap présente deux regroupements : l’un robuste et l’autre présentant deux sous-groupes plus forts (figure 84a). 89 LVnf ▼ DIVca ■ CUD ▼ VHcha ▼ PEY LZnf ROc2 53 91 25 Le premier ensemble est constitué par les groupes tardifs (DIVca, CUD, VHcha, PEY). Le second mêle majoritairement les échantillons ibériques et français les plus anciens. 100 71 52 8 BOI ▼ VILca ▼ LAnf/ca ▼ DIVcha ROc5 58 33 L’analyse en composantes principales reproduit à nouveau les éléments mis en évidence par les autres analyses. En effet, le nuage de points présente le regroupement des ensembles postérieurs, la position plus éclatée des échantillons anciens et le comportement mixte du Campaniforme (figure 84c). VILind 30 Le nuage de points produit par le multidimensional scaling complété par un minimal spanning tree reflète les résultats du dendrogramme (figure 84b). La cohérence d’une majorité des groupes tardifs ibériques et français est à nouveau visible. Les ensembles antérieurs semblent plus variables. Les Campaniformes se rattachent tantôt à l’homogénéité du Chalcolithique et du Bronze ancien, tantôt à la diversité des groupes anciens. -a- 0 0 38 79 -bROc2 0.24 VILind LZnf PEY ▼ BOI ▼ ▼ 0 ROc5 CUD ■ VHcha ▼DIVca VILca LVnf ▼ DIVcha Que dire de ces résultats ? La situation mise en évidence dans ce domaine est intéressante puisqu’on retrouve des différences plus marquées pour les périodes antérieures et une certaine harmonie des populations les plus récentes traduisant probablement des échanges plus marqués entre elles. Le Campaniforme au sein de cette configuration se situe entre ces deux courants. ▼ LAnf/ca -0.32 -0.4 0 0.3 -c- ROc2 LZnf 0.3 VILind VHcha ▼ PEY 0 Figure 84 : Résultats des analyses multivariées pour le domaine méridional. a) Analyse ascendante hiérarchique, selon la méthode de Ward, complétée par un bootstrap . b) Multidimensional scaling complété par un minimal spanning tree (stress=0.1379). c) Analyse en composantes principales (axe 1=67.4% et axe 2=22.8%). ▼■ CUD ▼ DIVcha VILca ▼ BOI LVnf ROc5 ▼ DIVca -0.5 ▼ LAnf/ca -0.48 0 0.64 183 Chapitre 7 7.2.3. A l’articulation entre les deux domaines : la Suisse 7.2.3.1. La Suisse La Suisse se situe à l’articulation entre les domaines méridional et oriental. Les vestiges sont inégalement répartis sur l’ensemble du territoire helvétique et présentent la dichotomie des traditions des différentes aires culturelles est-ouest. 7.2.3.1.1. La constitution des groupes testés Les populations se composent de plus de 520 individus et de 8 sites provenant essentiellement de la partie occidentale de la Suisse (figure 85). Elle présente une séquence sans hiatus important entre le 5e et le 3e millénaire, ainsi les échantillons se situent chronologiquement entre le Néolithique moyen et le Bronze ancien. - les ensembles retenus pour le Néolithique moyen Néolithique moyen de Sion (38 individus), sites n° 5, 8 : SCR Barmaz I (51 individus), site n° 1 : BAI Barmaz II (22 individus), site n° 2 : BAII Chamblandes (74 individus), site n° 3 : CHA Corseaux (44 individus), site n° 6 : COR - les ensembles retenus pour le Néolithique récent Petit-Chasseur, dolmen MXII (120 individus), sites n° 9 : MXII Petit-Chasseur, dolmen MVI (40 individus), sites n° 7 : MVIn - les ensembles retenus pour le Campaniforme Figure 85 : La Suisse, rappel de la localisation des sites (c.f. figure 13). Aesch (50 individus), sites n° 4 : AES Petit-Chasseur - Campaniforme (23 individus), sites n° 7 : PCcam Petit-Chasseur - fosse à incinération, mélanges Néolithique récent/Campaniforme (26 individus), sites n°7 : PCFI - les ensembles retenus pour le Bronze ancien Gisements appartenant au Bronze ancien valaisan (32 individus), sites n° 1, 7 : VSbz Toutes les observations sur les dents déciduales ont été regroupées selon la période à laquelle elles appartiennent : Néolithique moyen, Néolithique récent, Campaniforme et Bronze ancien. 7.2.3.1.2. Les incidences populationnelles Les incidences et les décomptes pour chaque variable se trouvent en table 45 pour la dentition permanente et en table 46 pour la dentition déciduale. La fosse à incinération du site de Petit-Chasseur à Sion (PCFI) ne comporte pas suffisamment d’observations, nous décidons de ne pas l’inclure au sein des analyses. Les autres groupes possèdent, pour la plupart, assez d’observations pour être intégrés. Lorsque les données sont insuffisantes pour l’un de ces ensembles, nous éliminons de préférence la variable. Pour la dentition permanente, les traits observés sur l’ensemble des membres d’un district présentent, en général, des fréquences d’expression similaires (table 45). Comme de coutume, les traits s’exprimant sur les racines et appartenant au système FU-DTS révèlent une importante constance d’expression. 184 L’analyse des données Les traits observés sur la dentition déciduale, malgré de petits effectifs, présentent des fréquences semblables pour les différentes périodes étudiées (Néolithique moyen, Néolithique récent, Campaniforme et Bronze ancien) (table 46). Seules les secondes molaires supérieures présentent quelques variations qui ne révèlent aucune différence flagrante entre les cultures. Les fréquences ajustées par la méthode de l’expression count E.C. pour chaque trait observé sur la dentition permanente sont données en table 47. DENTS PERMANENTES Maxillaire 1 Absence congénitale 2 Nombre de radicules 3 Racine appendiciforme 4 Nombre de racines des prémolaires 5 Nombre de racines des molaires 6 Racine de Carabelli 7 Racine paramolaire 8 Racine idiopathique 9 Degré de rotation 10 Convexité vestibulaire 11 Sillon traversant 12 Tubercule distal 13 Cingulum vestibulaire 14 Gouttière vestibulaire 15 Crêtes linguales marginales 16 Tubercule dentaire 17 Crêtes vestibulaires marginales 18 Crête mésiale 19 Crête accessoire distale 20 Cuspides accessoires 21 Odontome 22 Extension de l'émail 23 Parastyle 24 Trait de Carabelli 25 Métacone 26 Hypocone 27 Métaconule 28 Tubercule distal accessoire 29 Tubercule mésial paracone 30 Tubercule mésial accessoire 31 Protoconule 32 Tubercule lingual paracone M3 M2 M1 P2 P1 C ● I2 I1 ● ● ● ● ● ● ● Mandibule ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● 33 Absence congénitale 34 Nombre de radicules 35 Racine appendiciforme 36 Nombre racines canines 37 Racine de Tomes 38 Nombre racines molaires 39 Racine de Citroen 40 Racine paramolaire 41 Racine entomolaire 42 Racine idiopathique 43 Crêtes linguales marginales 44 Crêtes vestibulaires marginales 45 Crête accessoire distale 46 Odontome 47 Variation des cuspides linguales 48 Pli cuspide mésiolinguale 49 Fosse précuspidale 50 Crête trigonide intermédiaire 51 Crête trigonide distale 52 Nombre de cuspides 53 Dessin du sillon 54 Protostylide 55-56 Hypoconulide-Entoconulide 57 Métaconulide 58 Tubercule paracone 59 Tubercule de Citroen M3 M2 M1 P2 P1 C I2 I1 ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● Figure 86 : Représentation synthétique des dents clés retenues pour les analyses de la Suisse (n=49). 7.2.3.1.3. Les variables mobilisées dans les analyses Pour la Suisse, 49 variables sur les 79 traits localisés sur les dents clés ont été retenues. 27 d’entre elles s’observent sur le maxillaire et 22 sur la mandibule (figure 86). Les fréquences E.C. sont données en table 48. Les traits localisés sur les couronnes et sur les molaires sont, en général, mieux représentés. Les profils d’expression des variables observées sur les dents clés mobilisables présentent dans les grandes lignes des tendances similaires (figure 87). Il est tout de même possible de classer des spécificités selon les ensembles. Une même allure des profils appartenant aux ensembles datant du Néolithique moyen peut être mise en évidence. Ces groupes possèdent des fréquences élevées pour le nombre de racines des molaires supérieures, le métacone - T25 -, l’hypocone - T26 - et le nombre de racines des molaires inférieures T38, situation moins évidente pour le site de Chamblandes (CHA). Dès le Néolithique récent, les fréquences diffèrent selon les échantillons. Nous aurions pu imaginer qu’une même période favoriserait un profil similaire, ce qui n’est pas le cas. En effet, la séquence du dolmen MXII du Néolithique récent du Petit-Chasseur est proche du dolmen campaniforme de Aesch (AES). Les Campaniformes du Petit-Chasseur (PCcam) présentent un profil intermédiaire, tandis que le dolmen MVI se distingue complètement de tous ces ensembles. Enfin, le Bronze ancien (VSbz) se rattache au Néolithique moyen. Profils d’expression des différents groupes définis pour la Suisse 185 186 T31M1 T27M1 T24M1 T16I1 T11I1 T4P1 T1M3 T54M1 T37P1 T28M1 T19C T58M1 T16C T31M1 T27M1 T24M1 T16I1 T11I1 T4P1 T1M3 T54M1 T37P1 T28M1 T19C T1I2 T58M1 T31M1 T27M1 T24M1 T16I1 T11I1 T4P1 T1M3 T54M1 T37P1 T28M1 T19C T1I2 T58M1 0.70 T33M3 T33M3 T33M3 0.60 T34P1 T34P1 T34P1 0.50 T34I2 T34I2 T34I2 0.40 T47P2 T47P2 T47P2 0.30 T53M1 T53M1 T53M1 0.20 T34C T34C T34C 0.10 T2P1 T2P1 T2P1 0.70 T10I1 T10I1 T10I1 0.60 T55-56M1 T55-56M1 T55-56M1 0.50 T2M1 T2M1 T2M1 0.40 T52M1 T52M1 T52M1 T17P1 T45C T17P1 T45C T23M1 T17P1 T45C T23M1 T46P2 T44I2 T41M1 T36C T51M1 T46P2 T44I2 T43I2 T41M1 T36C T35I1 T51M1 T46P2 T44I2 T43I2 T41M1 T36C T35I1 T14C T14C T2I1 T14C T2I1 0.00 T17C T17C T17C 0.00 T2I1 T17I1 T17I1 T17I1 0.10 0.20 0.30 0.40 0.50 0.60 0.70 0.00 T2I1 T14C T17C T17I1 T22M1 T22P1 T22P1 T22M1 T22P1 T22M1 T22P1 T22M1 T35I1 T36C T41M1 T43I2 T44I2 T46P2 T51M1 T57M1 T35I1 T43I2 T51M1 T2C T57M1 T2C T57M1 T2C T2C T45C T17P1 T13I1 T16C T58M1 T1I2 T19C T28M1 T37P1 T54M1 T1M3 T4P1 T11I1 T16I1 T24M1 T27M1 T31M1 T33M3 T34P1 T34I2 T47P2 T53M1 T34C T2P1 T10I1 T55-56M1 T2M1 T52M1 T34M1 T38M1 T5M1 T25M1 T26M1 T23M1 T57M1 0.00 BAII BAII T23M1 T13I1 T16C T13I1 T16C T13I1 0.30 T34M1 T34M1 T34M1 0.20 T38M1 T38M1 T38M1 0.10 T5M1 T5M1 T5M1 T1I2 T25M1 T25M1 T26M1 T26M1 T25M1 BAI BAI T26M1 SRC SRC 0.10 0.20 0.30 0.40 CHA CHA 0.50 0.60 0.70 Chapitre 7 T19C T1I2 T58M1 T19C T1I2 T58M1 0.70 T28M1 T28M1 0.60 T37P1 T37P1 0.50 T54M1 T54M1 T14C T2I1 T14C T2I1 0.00 T17C 0.00 T17I1 T17C T36C T35I1 T36C T35I1 T17I1 T22P1 T41M1 T41M1 T22P1 T43I2 T43I2 T22M1 T44I2 T44I2 T22P1 T36C T35I1 T46P2 T46P2 T22M1 T41M1 T51M1 T51M1 T2C 0.00 T2I1 T14C T17C T17I1 T22M1 T43I2 T44I2 T46P2 T51M1 T57M1 T2C T57M1 T2C T23M1 T45C T17P1 T13I1 T1I2 T19C T28M1 T37P1 T54M1 T1M3 T4P1 T11I1 T16I1 T24M1 T27M1 T31M1 T33M3 T34P1 T34I2 T47P2 T53M1 T34C T2P1 T10I1 T55-56M1 T2M1 T52M1 T34M1 T38M1 T5M1 T57M1 0.40 T1M3 T1M3 0.30 T4P1 T4P1 0.20 T11I1 T11I1 0.10 T16I1 T16I1 0.70 T24M1 T24M1 0.60 T27M1 T27M1 0.50 T31M1 T31M1 0.40 T33M3 T33M3 0.30 T34P1 T34P1 0.20 T34I2 T34I2 0.10 T47P2 T47P2 T23M1 T53M1 T53M1 T23M1 T34C T34C T45C T2P1 T2P1 T17P1 T10I1 T10I1 T45C T55-56M1 T55-56M1 T17P1 T16C T2M1 T2M1 T16C T52M1 T52M1 T13I1 T34M1 T34M1 T16C T38M1 T38M1 T13I1 T58M1 T5M1 T5M1 T25M1 T25M1 T26M1 T26M1 T25M1 MXII MXII T26M1 COR COR 0.10 0.20 0.30 0.40 MVIn MVIn 0.50 0.60 0.70 0.00 T2I1 T14C T17C T17I1 T22M1 T22P1 T35I1 T36C T41M1 T43I2 T44I2 T46P2 T51M1 T57M1 T2C T23M1 T45C T17P1 T13I1 T16C T58M1 T1I2 T19C T28M1 T37P1 T54M1 T1M3 T4P1 T11I1 T16I1 T24M1 T27M1 T31M1 T33M3 T34P1 T34I2 T47P2 T53M1 T34C T2P1 T10I1 T55-56M1 T2M1 T52M1 T34M1 T38M1 T5M1 T25M1 T26M1 0.10 0.20 0.30 AES AES 0.40 0.50 0.60 0.70 L’analyse des données 187 188 0.20 0.40 0.50 0.60 0.70 T11I1 T11I1 T43I2 T41M1 T43I2 T41M1 Figure 87 : Profil pour chaque groupe testé dans les analyses, élaboré à partir des expression count des dents clés retenues pour la Suisse (n=49). Liste des variables selon l’ordre dans le profil, de haut en bas: T26M1-T25M1-T5M1-T38M1-T34M1-T2M1-T52M1-T2P1-T10I1-T55-56M1-T34C-T53M1-T47P2-T1M3-T4P1T11I1-T16I1-T24M1-T27M1-T31M1-T33M3-T34P1-T34I2-T1I2-T19C-T28M1-T37P1-T54M1-T13I1-T16CT58M1-T17P1-T2C-T23M1-T45C-T2I1-T14C-T17C-T17I1-T22M1-T22P1-T35I1-T36C-T41M1-T43I2-T44I2T46P2-T51M1-T57M1 T17C T14C T2I1 T17C T14C T2I1 0.00 T17I1 T17I1 0.00 T22P1 T22M1 T22P1 T22M1 T36C T44I2 T44I2 T35I1 T46P2 T46P2 T36C T51M1 T51M1 T35I1 T2C T57M1 T23M1 T2C T45C T23M1 T45C T57M1 T17P1 T17P1 T58M1 T16C T1I2 T13I1 T1I2 T58M1 T19C T16C T19C T28M1 T13I1 T37P1 T28M1 T37P1 T54M1 T16I1 T16I1 T54M1 T24M1 T24M1 T4P1 T27M1 T27M1 T1M3 T31M1 T31M1 T1M3 T33M3 T33M3 T4P1 T34I2 T34P1 T47P2 T47P2 T34P1 T53M1 T53M1 T34I2 T2P1 T34C T34C T10I1 T10I1 T2P1 T2M1 T52M1 T55-56M1 T52M1 T34M1 T55-56M1 T34M1 T38M1 T2M1 T5M1 T38M1 T5M1 0.30 T25M1 T25M1 0.10 T26M1 T26M1 PCcam PCcam 0.10 0.20 0.30 0.40 VSbz VSbz 0.50 0.60 0.70 Chapitre 7 L’analyse des données 7.2.3.1.4. Les analyses Les résultats des analyses multivariées montrent des tendances identiques (figure 88). -1 BAII 73 L’analyse ascendante hiérarchique complétée par un bootstrap présente deux regroupements forts où il est tout de même possible de distinguer trois sous-ensembles (figure 88a). COR 60 BAI 100 MVIn 64 ■ 22 8 CHA 47 ▼ 35 PCcam MXII 14 ▼ Le second mêle les échantillons Néolithique moyen de la région sédunoise (SRC) et l’ensemble tardif du Bronze ancien valaisan (VSbz). Le dolmen MVI datant du Néolithique récent se rattache aux deux derniers ensembles (le groupe réunissant Néolithique moyen et Bronze ancien et le groupe avec les populations récentes), en gardant tout de même une certaine distance. VSbz SRC 52 Le premier rassemble uniquement des populations du Néolithique moyen, soit les deux nécropoles de Barmaz (BAI et BAII) et le site de Corseaux (COR). Le dernier réunit les ensembles campaniformes (AES et PCcam) et l’un des deux dolmens du Néolithique récent (MXII). -a- 0 0 AES -b0.24 ■ VSbz SRC BAI BAII 0 COR CHA MVIn ▼PCcam MXII Nous retrouvons une même configuration dans la représentation issue du multidimensional scaling complété par un minimal spanning tree (figure 88b). ▼AES -0.3 Le nuage de points présente une tendance similaire, avec une certaine cohésion des populations du Néolithique moyen et des groupements plus éclatés pour les périodes postérieures. L’analyse en composantes principales reproduit les éléments mis en évidence par les autres analyses. En effet, la représentation montre à nouveau le regroupement des ensembles du Néolithique moyen et la position plus éclatée du Campaniforme (AES et PCcam), du Néolithique récent (MXII et MVI) et du Bronze ancien (VSbz) (figure 88c). -0.6 0 -cMVIn 0.24 BAII 0 COR Nous devrions, lors des deux prochaines analyses concernant la position de la Suisse face aux différents domaines étudiés, pouvoir préciser cette situation. BAI SRC ■ VSbz ▼PCcam CHA Que dire de ces résultats ? Les différentes analyses s’accordent pour défendre l’idée d’une franche cohésion des populations du Néolithique moyen en territoire helvétique et d’une certaine variabilité des ensembles tardifs qui traduirait probablement une mobilité ou un apport populationnel modéré pendant ces différentes périodes. 0.48 MXII AES ▼ -0.24 -0.8 0 Figure 88 : Résultats des multivariées pour la Suisse. 0.6 analyses a) Analyse ascendante hiérarchique, selon la méthode de Ward, complétée par un bootstrap . b) Multidimensional scaling complété par un minimal spanning tree (stress=0.08484). c) Analyse en composantes principales (axe 1=50.9% et axe 2=14.9%). 189 Chapitre 7 7.2.3.2. Les relations de la Suisse avec le domaine oriental Cette partie de l’analyse vise à établir les relations entre les échantillons suisses et les groupes orientaux Bohême et Hongrie -. 7.2.3.2.1. La constitution des groupes testés Nous avons inclus l’ensemble des groupes intégrés lors des analyses intrarégionales pour le domaine oriental. Seuls les ensembles tardifs helvétiques sont sélectionnés, soit le Néolithique récent, le Campaniforme et le Bronze ancien. Pour rappel, les ensembles retenus pour la Suisse : - Néolithique récent - Bronze ancien Petit-Chasseur, dolmen MXII (120 individus), sites n° 9 : MXII Gisements appartenant au Bronze ancien valaisan (32 individus), sites n° 1, 7 : VSbz Petit-Chasseur, dolmen MVI (40 individus), sites n° 7 : MVIn - Campaniforme Aesch (50 individus), sites n° 4 : AES Petit-Chasseur - Campaniforme (23 individus), sites n° 7 : PCcam Pour rappel, les échantillons sélectionnés pour la Bohême : - Cordé Vikletice (90 individus), site n° 60 : VIK Bohême centrale (40 individus), sites n° 3, 16, 32, 36, 56, 59 : BCca Mochov (21 individus), site n° 25 : MOC Bohême occidentale (58 individus), sites n° 1, 2, 8, 9, 10, 12, 14, 17, 19, 26, 27, 28, 40, 41, 42, 47, 49, 50, 51, 53, 55, 63 : BWco - Unetice Bohême orientale (36 individus), sites n° 5, 7 : BEco Bohême occidentale (38 individus), sites n° 2, 8, 22, 44, 45, 53, 58, 60, 61: BWun Bohême centrale (24 individus), sites n° 4, 11, 15, 23, 29, 35, 37, 52, 54 : BCco Brezno (38 individus), site n° 5 : BRE - Campaniforme Bohême occidentale (20 individus), sites n° 18, 20, 42, 43, 44, 46, 48, 53, 62 : BWca Bohême orientale (41 individus), sites n° 6, 13, 24, 39 : BEun Bohême centrale (36 individus), sites n° 23, 31, 33, 34, 35, 37, 38, 57 : BCun Mala Ohrada (47 individus), site n° 30 : MaOr Bohême orientale (17 individus), sites n° 5, 7, 21, 39 : BEca Pour rappel, les 4 groupes culturels hongrois : 190 Gisements - Campaniforme (19 individus), sites n° 1, 2, 3, 13 : CAM Gisements - Obedà-Pitvaros (10 individus), sites n° 6, 8, 12 : OB-PIT Gisements - Nagyrev (35 individus), sites n° 4, 5, 10, 11 : NAG Gisements - Perjamos (26 individus), sites n° 9, 14 : PER L’analyse des données DENTS PERMANENTES Maxillaire 1 Absence congénitale 2 Nombre de radicules 3 Racine appendiciforme 4 Nombre de racines des prémolaires 5 Nombre de racines des molaires 6 Racine de Carabelli 7 Racine paramolaire 8 Racine idiopathique 9 Degré de rotation 10 Convexité vestibulaire 11 Sillon traversant 12 Tubercule distal 13 Cingulum vestibulaire 14 Gouttière vestibulaire 15 Crêtes linguales marginales 16 Tubercule dentaire 17 Crêtes vestibulaires marginales 18 Crête mésiale 19 Crête accessoire distale 20 Cuspides accessoires 21 Odontome 22 Extension de l'émail 23 Parastyle 24 Trait de Carabelli 25 Métacone 26 Hypocone 27 Métaconule 28 Tubercule distal accessoire 29 Tubercule mésial paracone 30 Tubercule mésial accessoire 31 Protoconule 32 Tubercule lingual paracone M3 M2 M1 P2 P1 ● C ● ● I1 ● ● ● ● Mandibule ● ● ● ● ● ● ● ● ● ● ● I2 ● 33 Absence congénitale 34 Nombre de radicules 35 Racine appendiciforme 36 Nombre racines canines 37 Racine de Tomes 38 Nombre racines molaires 39 Racine de Citroen 40 Racine paramolaire 41 Racine entomolaire 42 Racine idiopathique 43 Crêtes linguales marginales 44 Crêtes vestibulaires marginales 45 Crête accessoire distale 46 Odontome 47 Variation des cuspides linguales 48 Pli cuspide mésiolinguale 49 Fosse précuspidale 50 Crête trigonide intermédiaire 51 Crête trigonide distale 52 Nombre de cuspides 53 Dessin du sillon 54 Protostylide 55-56 Hypoconulide-Entoconulide 57 Métaconulide 58 Tubercule paracone 59 Tubercule de Citroen M3 M2 M1 P2 P1 C I2 I1 ● ● ● ● ● ● ● ● ● ● ● Figure 89 : Représentation synthétique des dents clés retenues pour les analyses Suisse et domaine oriental (n=30). 7.2.3.2.2. Les variables mobilisées dans les analyses Pour cette partie de l’étude, 30 variables sur les 79 traits localisés sur les dents clés ont été retenues. 19 d’entre elles s’observent sur le maxillaire et 11 sur la mandibule (figure 89). Les fréquences E.C. sont données table 49. Les traits localisés sur les couronnes et sur les molaires sont, en général, mieux représentés. 191 Chapitre 7 7.2.3.2.3. Les analyses Les résultats des analyses multivariées sont très intéressants et montrent des configurations comparables (figure 90). -1 28 8 99 100 25 33 58 83 16 12 14 BEco ■ BCun ■ BRE ■ BEun ■ MaOr OB-PIT BCco ▼ BCca 21 25 3 ▼ PCcam ▼ CAM ▼ BWca 10 10 MVIn ■ BWun MXII ■ VSbz 52 Le premier est composé de la totalité des sites helvétiques qui ne se mêlent pas aux ensembles orientaux. Au sein de cette configuration, deux groupes se mêlent aux populations suisses, soit le Bronze ancien de Bohême occidentale (BCun) et les Campaniformes hongrois (CAM). Un second groupe est constitué par le Bronze ancien tchèque (BCun, BRE, BCun et MaOr). A l’intérieur, les Campaniformes occidentaux (BWca) et les Cordés orientaux (BEco) se rattachent aux ensembles unetices. ▼ AES 50 28 L’analyse ascendante hiérarchique complétée par un bootstrap présente des associations, entre populations, peu robustes pour les grandes dichotomies (figure 90a). Nous distinguons tout de même des regroupements importants. -a- 0 0 20 22 VIK BWco ▼ NAG PER ▼ BEca ▼ MOC 11 -bBWca 0.16 ▼ BCco Un dernier regroupement peu robuste est formé par les Cordés et les Campaniformes tchèques et par les différentes périodes hongroises (à l’exception du Campaniforme). VSbz ■ Les autres ensembles hongrois se distinguent Campaniforme régional et se situent légèrement à l’écart. Que dire de ces résultats ? Au sein de cette superposition de cultures, périodes et territoires, nous constatons les mêmes tendances distinguées lors des analyses régionales. Les éléments importants à retenir : les sites suisses ne se mêlent pas au domaine oriental et les Campaniformes hongrois semblent bien plus proches de leur contemporains occidentaux. 192 AES ▼ BWco ■ MXIIMaOr VIK ▼ PCcam ▼ PER OB-PIT ▼ MVIn MOC -0.2 -0.4 0 0.4 -cOB-PIT 0.24 MVIn du L’analyse en composantes principales reproduit à nouveau les éléments mis en évidence par les autres analyses. On retrouve la concentration des sites suisses, celle du Bronze ancien tchèque, la variabilité cordée et campaniforme ainsi que l’éloignement du Campaniforme hongrois de ses congénères (figure 90c). BCun BEco BRE ■ BEun ▼ ▼ NAG ■ BCca BEca BWun ■ 0 Les sites helvétiques se regroupent également, à l’exception du Bronze ancien (VSbz) qui s’associe au Campaniforme hongrois (CAM). ▼ ■ Nous retrouvons la même configuration au sein du nuage de points issu du multidimensional scaling complété par un minimal spanning tree (figure 90b). Le Bronze ancien tchèque montre à nouveau une concentration homogène. Les ensembles cordés et campaniformes tchèques, au contraire, présentent une certaine hétérogénéité. CAM VSbz ■ AES▼ 0 MXII ▼ ■ PCcam BRE BEco -0.24 -0.8 VIK NAG ■ ▼PER ▼BEca ■ ■ BEun MaOr ▼ ▼ MOC ▼ CAM ■ BCca BCun BWun BWco BCco ▼BWca 0 0.6 Figure 90 : Résultats des analyses multivariées Suisse et domaine oriental. a) Analyse ascendante hiérarchique, selon la méthode de Ward, complétée par un bootstrap . b) Multidimensional scaling complété par un minimal spanning tree (stress=0.07759). c) Analyse en composantes principales (axe 1=80.2% et axe 2=6.7%). L’analyse des données 7.2.3.3. Les relations de la Suisse avec le domaine méridional Cette partie de l’analyse vise à établir les relations entre les échantillons suisses et les groupes méridionaux - sud de la France et nord de l’Espagne -. 7.2.3.3.1. La constitution des groupes testés Nous avons inclus l’ensemble des groupes intégrés lors des analyses intrarégionales pour le domaine méridional. Seuls les ensembles tardifs helvétiques sont sélectionnés, soit le Néolithique récent, le Campaniforme et le Bronze ancien. Pour rappel, les ensembles retenus pour la Suisse : - Néolithique récent - Bronze ancien Petit-Chasseur, dolmen MXII (120 individus), sites n° 9 : MXII Gisements appartenant au Bronze ancien valaisan (32 individus), sites n° 1, 7 : VSbz Petit-Chasseur, dolmen MVI (40 individus), sites n° 7 : MVIn - Campaniforme Aesch (50 individus), sites n° 4 : AES Petit-Chasseur - Campaniforme (23 individus), sites n° 7 : PCcam Pour rappel, les ensembles retenus pour le sud de la France : Boileau - Néolithique final (130 individus), site n° 4 : BOI Villedubert - Campaniforme (70 individus), site n° 1 : VILca Peyraoutes - Chalcolithique/Campaniforme (172 individus), site n° 2 : PEY Villedubert - phase indéterminée (19 individus), site n° 1 : VILind Roaix, couche 5 - Chalcolithique (92 individus), site n° 3 : ROc5 Cudières - Chalcolithique/Bronze ancien (98 individus), site n° 5 : CUD Roaix, couche 2 - Chalcolithique (49 individus), site n° 3 : ROc2 Pour rappel, les échantillons sélectionnés pour le nord de l’Espagne : La Velilla - Néolithique final (50 individus), site n° 9 : LVnf Gisements - Chalcolithique (13 individus), sites n° 2, 6, 11, 14 : DIVcha Los Zumacalles - Néolithique final (18 individus), site n° 12 : LZnf Valle de las Higueras - Chalcolithique (38 individus), site n° 13 : VHcha Las Arnillas - Néolithique final/Campaniforme (24 individus), site n°10 : LAnf/ca Gisements - Campaniforme (18 individus), sites n° 3, 7, 8, 13 : DIVca 193 Chapitre 7 DENTS PERMANENTES Maxillaire 1 Absence congénitale 2 Nombre de radicules 3 Racine appendiciforme 4 Nombre de racines des prémolaires 5 Nombre de racines des molaires 6 Racine de Carabelli 7 Racine paramolaire 8 Racine idiopathique 9 Degré de rotation 10 Convexité vestibulaire 11 Sillon traversant 12 Tubercule distal 13 Cingulum vestibulaire 14 Gouttière vestibulaire 15 Crêtes linguales marginales 16 Tubercule dentaire 17 Crêtes vestibulaires marginales 18 Crête mésiale 19 Crête accessoire distale 20 Cuspides accessoires 21 Odontome 22 Extension de l'émail 23 Parastyle 24 Trait de Carabelli 25 Métacone 26 Hypocone 27 Métaconule 28 Tubercule distal accessoire 29 Tubercule mésial paracone 30 Tubercule mésial accessoire 31 Protoconule 32 Tubercule lingual paracone M3 M2 M1 P2 P1 ● C ● ● I1 ● ● ● ● Mandibule ● ● ● ● ● ● ● ● ● ● ● I2 ● 33 Absence congénitale 34 Nombre de radicules 35 Racine appendiciforme 36 Nombre racines canines 37 Racine de Tomes 38 Nombre racines molaires 39 Racine de Citroen 40 Racine paramolaire 41 Racine entomolaire 42 Racine idiopathique 43 Crêtes linguales marginales 44 Crêtes vestibulaires marginales 45 Crête accessoire distale 46 Odontome 47 Variation des cuspides linguales 48 Pli cuspide mésiolinguale 49 Fosse précuspidale 50 Crête trigonide intermédiaire 51 Crête trigonide distale 52 Nombre de cuspides 53 Dessin du sillon 54 Protostylide 55-56 Hypoconulide-Entoconulide 57 Métaconulide 58 Tubercule paracone 59 Tubercule de Citroen M3 M2 M1 P2 P1 C I2 I1 ● ● ● ● ● ● ● ● ● ● ● Figure 91 : Représentation synthétique des dents clés retenues pour les analyses Suisse et domaine méridional (n=30). 7.2.3.3.2. Les variables mobilisées dans les analyses Pour cette fraction, 30 variables sur les 79 traits localisés sur les dents clés ont été retenues. 19 d’entre elles s’observent sur le maxillaire et 11 sur la mandibule (figure 91). Les fréquences E.C. sont données table 50. Les traits localisés sur les couronnes et sur les molaires sont, en général, mieux représentés. 194 L’analyse des données 7.2.3.3.3. Les analyses Les résultats des analyses comparables (figure 92). multivariées sont forts -1 Le second est constitué par l’ensemble des échantillons suisses et de la majorité des groupes récents ibériques et français (PEY, CUD, DIVca, LAnf). Les sites espagnols de la Velilla (LVnf) et de la Valle de las Higueras (VHcha) se joignent à ce regroupement. La représentation issue du multidimensional scaling complété par un minimal spanning tree propose une configuration similaire (figure 92b). Les ensembles tardifs ibériques et français ainsi que les groupes suisses présentent à nouveau une concentration homogène. Le Néolithique final semble encore une fois plus variable. L’unique échantillon campaniforme s’isolant des périodes tardives est encore le site de Villedubert (VILca). 71 54 31 59 ▼ 100 70 40 10 ▼ 92 ▼ ■ ▼ ▼ 65 43 35 26 29 ▼ 52 22 ■ ▼ DIVcha ROc5 LZnf ROc2 VILind BOI VILca LVnf MXII PCcam LAnf/ca MVIn VSbz VHcha PEY DIVca CUD AES -b0.24 ROc2 VILind VSbz LZnf ■ 0 ▼ ROc5 VHcha ▼ ▼PEY CUD ■ DIVca BOI VILca ▼ AES ▼ MXII ▼ LVnf PCcam MVIn ▼ DIVcha ▼ LAnf/ca -0.3 -0.68 L’analyse en composantes principales met en avant les éléments constatés lors des analyses précédentes. En effet, le nuage de points présente le regroupement des ensembles récents, la position plus éclatée du Néolithique final et du Chalcolithique (figure 92c). ▼ 79 69 L’analyse ascendante hiérarchique complétée par un bootstrap présente deux regroupements forts (figure 92a). Le premier est composé par les échantillons les plus anciens français (ROc5, ROc2 et BOI) et l’ensemble indéterminé de Villedubert (VILind), par le site de Los Zumacalles (LZnf) et le groupe chalcolithique pour la péninsule Ibérique. Le seul ensemble campaniforme présent est l’ensemble de Villedubert (VILca). -a- 0 0 0 0.24 -cROc2 LZnf 0.3 VSbz ▼ ■ VILind VHcha ▼ PEY CUD■ ▼ DIVca Que dire de ces résultats ? 0 Les échantillons helvétiques se regroupent de manière distincte sur l’ensemble des configurations. Ils se rattachent aux périodes récentes ibériques et françaises. BOI ▼ AES▼ MVIn VILca ▼PCcam LVnf MXII ROc5 ▼ DIVcha Les groupes anciens du domaine méridional présentent une plus grande variabilité. Cet élément s’expliquerait probablement par une mobilité ou des échanges populationnels moindres pendant ces périodes. Ceci ne semble pas être le cas en territoire helvétique et pour les ensembles tardifs méridionaux où l’homogénéité apparente permettrait d’avancer une accentuation des influences externes. ▼ LAnf/ca -0.5 -0.48 0 0.64 Figure 92 : Résultats des analyses multivariées Suisse et domaine méridional. a) Analyse ascendante hiérarchique, selon la méthode de Ward, complétée par un bootstrap . b) Multidimensional scaling complété par un minimal spanning tree (stress=0.05041). c) Analyse en composantes principales (axe 1=70.2% et axe 2=19.8%). 195 Chapitre 7 7.2.4. Les relations entre les populations campaniformes Cette partie de l’analyse s’attache à déterminer les affinités entre les échantillons campaniformes des cinq régions étudiées. 7.2.4.1. La constitution des groupes testés Nous avons sélectionné uniquement les groupes campaniformes pour l’ensemble des régions. Les sites de Les Peyraoutes (sud de la France) et de Las Arnillas (nord de l’Espagne) présentent tous deux des échantillons culturellement mixtes. - Bohême - Nord de l’Espagne Bohême occidentale (20 individus), sites n° 18, 20, 42, 43, 44, 46, 48, 53, 62 : BWca Las Arnillas - Néolithique final/Campaniforme (24 individus), site n°10 : LAnf/ca Bohême orientale (17 individus), sites n° 5, 7, 21, 39 : BEca Gisements - Campaniforme (18 individus), sites n° 3, 7, 8, 13 : DIVca Bohême centrale (40 individus), sites n° 3, 16, 32, 36, 56, 59 : BCca - Suisse Mochov (21 individus), site n° 25 : MOC Aesch (50 individus), sites n° 4 : AES - Hongrie Petit-Chasseur - Campaniforme (23 individus), sites n° 7 : PCcam Gisements - Campaniforme (19 individus), sites n° 1, 2, 3, 13 : CAM - Sud de la France Peyraoutes - Chalcolithique/Campaniforme (172 individus), site n° 2 : PEY Villedubert - Campaniforme (70 individus), site n° 1 : VILca DENTS PERMANENTES Maxillaire 1 Absence congénitale 2 Nombre de radicules 3 Racine appendiciforme 4 Nombre de racines des prémolaires 5 Nombre de racines des molaires 6 Racine de Carabelli 7 Racine paramolaire 8 Racine idiopathique 9 Degré de rotation 10 Convexité vestibulaire 11 Sillon traversant 12 Tubercule distal 13 Cingulum vestibulaire 14 Gouttière vestibulaire 15 Crêtes linguales marginales 16 Tubercule dentaire 17 Crêtes vestibulaires marginales 18 Crête mésiale 19 Crête accessoire distale 20 Cuspides accessoires 21 Odontome 22 Extension de l'émail 23 Parastyle 24 Trait de Carabelli 25 Métacone 26 Hypocone 27 Métaconule 28 Tubercule distal accessoire 29 Tubercule mésial paracone 30 Tubercule mésial accessoire 31 Protoconule 32 Tubercule lingual paracone 196 M3 M2 M1 P2 P1 ● C ● ● I1 ● ● ● ● Mandibule ● ● ● ● ● ● ● ● ● ● ● I2 ● 33 Absence congénitale 34 Nombre de radicules 35 Racine appendiciforme 36 Nombre racines canines 37 Racine de Tomes 38 Nombre racines molaires 39 Racine de Citroen 40 Racine paramolaire 41 Racine entomolaire 42 Racine idiopathique 43 Crêtes linguales marginales 44 Crêtes vestibulaires marginales 45 Crête accessoire distale 46 Odontome 47 Variation des cuspides linguales 48 Pli cuspide mésiolinguale 49 Fosse précuspidale 50 Crête trigonide intermédiaire 51 Crête trigonide distale 52 Nombre de cuspides 53 Dessin du sillon 54 Protostylide 55-56 Hypoconulide-Entoconulide 57 Métaconulide 58 Tubercule paracone 59 Tubercule de Citroen M3 M2 M1 P2 P1 C I2 I1 ● ● ● ● ● ● ● ● ● ● ● Figure 93 : Représentation synthétique des dents clés retenues pour les analyses entre les ensembles campaniformes (n=30). L’analyse des données 7.2.4.2. Les variables mobilisées dans les analyses Pour cette partie, 30 variables sur les 79 traits localisés sur les dents clés ont été retenues. 19 d’entre elles s’observent sur le maxillaire et 11 sur la mandibule (figure 93). Les fréquences E.C. sont données en table 51. Les traits localisés sur les couronnes et sur les molaires sont, en général, mieux représentés. Les profils d’expression des variables observées sur les dents clés mobilisables sont très intéressants et présentent des séquences cohérentes où il est possible de distinguer des spécificités selon les domaines étudiés (figure 94). Les Campaniformes de Bohême possèdent des profils très proches avec des expressions élevées des variables qui ont pu être mobilisées pour les analyses. En particulier, quatre traits expriment des fréquences élevées, notamment le nombre de racines des molaires supérieures - T5 -, le métacone - T25 -, l’hypocone - T26 - et l’hypoconulide-entoconulide - T55-56 -. Les ensembles appartenant au domaine méridional expriment les mêmes variables, mais avec des incidences nettement moins prononcées. Au sein du domaine oriental, deux ensembles se particularisent. Le campaniforme hongrois (CAM) possède un profil comparable à ceux que l’on rencontre pour les groupes méridionaux avec des fréquences moins élevées. Les Campaniformes de la Bohême occidentale (BWca) révèlent également un profil plus modéré sur l’ensemble des variables, à l’exception de trois traits (métacone - T25 -, hypocone T26 - et nombre de radicules des prémolaires supérieures - T2 -) pour lesquels les incidences se rattachent au domaine méridional. Le profil de la phase campaniforme du site français de Villedubert (VILca), bien que présentant une séquence occidentale, possède des expressions franchement orientales pour deux variables (métacone T25 - et hypocone - T26 -). Le site ibérique de Las Arnillas (LAnf/ca), échantillon mixte Néolithique final et Campaniforme, se différencie de l’ensemble des groupes campaniformes par la fréquence élevée de trois variables (nombre de racines des molaires inférieures - T38 -, nombre de cuspides - T52 - et hypoconulide-entoconulide T55-56 -). Au-delà de ces traits, son profil reste fidèle au domaine méridional auquel il appartient. Nous avions déjà noté cette distinction au sein de l’analyse régionale. En effet, il s’y particularisait avec le groupe chalcolithique ibérique (DIVcha) par l’expression de ces mêmes traits. Profils d’expression des différents groupes définis pour l’analyse entre ensembles campaniformes 197 198 0.60 0.70 0.10 0.40 0.50 0.60 0.70 0.10 0.20 0.30 0.40 0.50 0.60 0.70 T57M1 T41M1 T23M1 T1I2 T1P2 T57M1 T46P2 T41M1 T23M1 T1I2 T1P2 T57M1 T46P2 T41M1 T23M1 T17C T17P1 T8M1 T7M1 T3C T1I2 T1P2 T1M3 T57M1 T46P2 T41M1 T23M1 T17C T17P1 T8M1 T7M1 T3C T1I2 T1P2 T1M3 0.00 T58M1 T58M1 T58M1 T58M1 0.00 T59M1 T59M1 T59M1 T59M1 0.00 T1M3 T3C T7M1 T8M1 T17P1 T17C T27M1 T27M1 T27M1 T27M1 0.00 T1M3 T3C T7M1 T8M1 T17P1 T17C T46P2 T2C T28M1 T2C T28M1 T2C T28M1 T2C T16C T16C T16C T16C T28M1 T2P1 T54M1 T2P1 T54M1 T2P1 T54M1 T2P1 T54M1 T47P2 T24M1 T47P2 T24M1 T47P2 T24M1 T47P2 0.30 T2M1 T34M1 T24M1 0.20 T34M1 T34M1 T34M1 0.50 T2M1 T2M1 T2M1 0.40 T38M1 T38M1 T38M1 0.30 T38M1 T52M1 T52M1 T52M1 0.20 T52M1 T55-56M1 T55-56M1 T55-56M1 0.10 T26M1 0.87 T26M1 T26M1 T26M1 T55-56M1 T5M1 T5M1 0.71 T25M1 0.92 0.82 T25M1 T5M1 0.72 T25M1 BEca TCHEQUIE - BEca T5M1 BWca TCHEQUIE - BWca T25M1 CAM HONGRIE - CAM 0.10 0.20 0.30 BCca 0.40 0.50 TCHEQUIE - BCca 0.60 0.70 0.75 0.78 Chapitre 7 0.60 0.10 0.20 0.40 0.50 0.60 0.70 0.10 0.20 0.30 0.40 0.50 0.60 0.70 T24M1 0.30 T24M1 T24M1 T59M1 T58M1 T57M1 T46P2 T41M1 T23M1 T17C T17P1 T8M1 T7M1 T3C T1I2 T1P2 T1M3 T27M1 T59M1 T58M1 T57M1 T46P2 T41M1 T23M1 T17C T17P1 T8M1 T7M1 T3C T1I2 T1P2 T1M3 T27M1 T59M1 T58M1 T57M1 T46P2 T41M1 T23M1 T17C T17P1 T8M1 T7M1 T3C T1I2 T1P2 T1M3 T28M1 T27M1 T59M1 T58M1 T57M1 T46P2 T41M1 T23M1 T17C T17P1 T8M1 T7M1 T3C T1I2 T1P2 T1M3 0.00 T27M1 T28M1 T28M1 0.00 T2C T28M1 T2C T2C T2C 0.00 T16C T16C T16C T16C 0.00 T2P1 T54M1 T2P1 T54M1 T2P1 T54M1 T2P1 T54M1 0.70 T47P2 T24M1 T47P2 T47P2 T47P2 0.50 T34M1 T34M1 T34M1 T34M1 0.40 T2M1 T2M1 T2M1 T2M1 0.30 T38M1 T38M1 T38M1 T38M1 0.20 T52M1 T52M1 T52M1 T52M1 T26M1 T55-56M1 T26M1 T55-56M1 T26M1 T26M1 T5M1 T25M1 T55-56M1 0.84 PCcam SUISSE - PCcam T55-56M1 T5M1 T5M1 0.76 T25M1 T25M1 0.83 T5M1 AES SUISSE - AES T25M1 0.10 MOC TCHEQUIE - MOC 0.10 0.20 0.30 0.40 0.50 PEY FRANCE - PEY 0.60 0.70 L’analyse des données 199 200 0.10 0.20 0.30 0.40 0.50 0.60 0.70 0.10 0.20 0.30 0.40 0.50 0.60 0.70 T23M1 T23M1 T23M1 Figure 94 : Profil pour chaque groupe testé dans les analyses, élaboré à partir des expression count des dents clés retenues pour les ensembles campaniformes (n=30). Liste des variables selon l’ordre dans le profil, de haut en bas: T25M1-T5M1-T26M1-T55-56M1-T52M1-T38M1-T2M1-T34M1-T47P2-T24M1-T2P1-T54M1-T16C-T2C-T28M1T27M1-T1M3-T1P2-T1I2-T3C-T7M1-T8M1-T17P1-T17C-T23M1-T41M1-T46P2-T57M1-T58M1-T59M1 T1P2 T1M3 T1P2 T1M3 0.00 T1P2 T1M3 T1I2 T3C T1I2 0.00 T3C T1I2 T3C 0.00 T7M1 T7M1 T8M1 T41M1 T41M1 T41M1 T8M1 T46P2 T46P2 T46P2 T7M1 T57M1 T57M1 T57M1 T8M1 T58M1 T58M1 T58M1 T17C T59M1 T59M1 T59M1 T17P1 T27M1 T27M1 T27M1 T17C T28M1 T28M1 T28M1 T17P1 T2C T2C T2C T17C T16C T16C T16C T17P1 T2P1 T54M1 T2P1 T54M1 T2P1 T54M1 T47P2 T24M1 T47P2 T24M1 T2M1 T34M1 T2M1 T34M1 T2M1 T34M1 T47P2 T38M1 T38M1 T38M1 T24M1 T52M1 T52M1 T52M1 T26M1 T26M1 T55-56M1 T5M1 T25M1 T5M1 T25M1 T55-56M1 0.71 0.72 LAnf/ca ESPAGNE - LAnf/ca T55-56M1 T26M1 T5M1 T25M1 VILca FRANCE - VILca 0.10 0.20 0.30 0.40 0.50 DIVca ESPAGNE - DIVca 0.60 0.70 Chapitre 7 L’analyse des données 7.2.4.3. Les analyses Les résultats des analyses multivariées configurations très intéressantes (figure 95). révèlent des -1 30 L’analyse ascendante hiérarchique complétée par un bootstrap présente deux regroupements qui ne sont pas vraiment robustes mais très révélateurs (figure 95a). 53 61 56 100 Le premier est composé par l’ensemble des groupes méridionaux (LA-ES, PEY-F et DIV-ES) et par les échantillons helvétiques (AES-CH et PC-CH). Le second est constitué par l’ensemble des échantillons campaniformes orientaux (BC-CZ, BW-CZ, BE-CZ, MOC-CZ, CAM-HU). Au sein de ce deuxième regroupement, le site de Villedubert (VIL-F), se situant géographiquement dans le domaine méridional, forme un groupe cohérent avec les Campaniformes tchèques occidentaux (BW-CZ). La position des Campaniformes hongrois (CAM-HU) se situe un peu en marge de la concentration orientale. -a- 0 0 6 46 91 25 PEY-F AES-CH DIV-ES CAM-HU 72 PC-CH 18 LA-ES BC-CZ VIL-F BW-CZ BE-CZ MOC-CZ 12 -b0.36 LA-ES Nous retrouvons les mêmes dispositions dans la représentation issue du multidimensional scaling complété par un minimal spanning tree (figure 95b). L’agglomération des groupes occidentaux (domaine méridional et Suisse) est à nouveau constatée. Seul l’ensemble ibérique de Las Arnillas (LA-ES) se détache un peu de cette concentration. Les Campaniformes orientaux se distinguent et forment un groupe un peu plus éclaté. DIV-ES AES-CH 0 MOC-CZ PC-CH VIL-F BW-CZ PEY-F BC-CZ BE-CZ CAM-HU -0.2 -0.4 Les Campaniformes hongrois (CAM-HU) et le gisement de Villeduvert (VIL-F) se situent en zone intermédiaire entre les deux grands courants. L’analyse en composantes principales reflète à nouveau les éléments mis en évidence lors des analyses précédentes. En effet, on distingue la concentration des ensembles méridionaux, le regroupement plus éclaté des orientaux et la position entre ces deux courants des hongrois et de Villedubert (figure 95c). Le site espagnol de Las Arnillas est un mélange d’occupations Néolithique final et Campaniforme. Cette condition expliquerait peut être son éloignement de la concentration des ensembles occidentaux. Nous devons noter la proximité de certains ensembles - les Campaniformes hongrois et le site de Villedubert - avec les aires géographiques opposées. LA-ES 0.4 PC-CH AES-CH BE-CZ DIV-ES Que dire de ces résultats ? D’après les représentations, les échanges et les influences externes semblent majeures dans l’aire occidentale. 0.5 -c- 0 Il est possible de mettre en évidence une dichotomie nette et franche des populations campaniformes du domaine oriental des échantillons méridionaux. Les Campaniformes helvétiques se rattachent sans équivoque à ce dernier domaine. 0 VIL-F PEY-F -0.24 CAM-HU -0.36 MOC-CZ BC-CZ BW-CZ 0 0.6 Figure 95 : Résultats des analyses multivariées pour les ensembles campaniformes. a) Analyse ascendante hiérarchique, selon la méthode de Ward, complétée par un bootstrap . b) Multidimensional scaling complété par un minimal spanning tree (stress=0.047). c) Analyse en composantes principales (axe 1=72.7% et axe 2=16.4%). 201 Chapitre 7 7.2.5. La Bohême : une analyse sexes séparés L’unique région présentant un nombre suffisant de déterminations sexuelles pour chaque période étudiée est la Bohême. Une analyse sexes séparés a alors été envisagée afin de déterminer si des différences peuvent être mises en évidence selon le sexe des individus dans une perspective de dynamique des peuplements. 7.2.5.1. La constitution des groupes testés Nous avons réuni les individus sexuellement déterminés - d’après les données anthropologiques - selon la période à laquelle ils appartiennent : Cordé-homme : individus cordés sexés masculins (67 individus) Cordé-femme : individus cordés sexés féminins (26 individus) Campa-homme : individus campaniformes sexés masculins (35 individus) Campa-femme : individus campaniformes sexés féminins (11 individus) Unet-homme : individus unetices sexés masculins (76 individus) Unet-femme : individus unetices sexés féminins (25 individus) Les indéterminés (sujets adultes et immatures) de chaque culture ont également été réunis. Nous les intégrerons dans les analyses uniquement dans une optique comparative : Cordé-indéter : individus cordés non sexés (114 individus) Campa-indéter : individus campaniformes non sexés (52 individus) Unet-indéter : individus unetices non sexés (100 individus) 7.2.5.2. Les incidences populationnelles Les incidences ont été calculées uniquement sur les dents clés de la dentition permanente. Les fréquences et les décomptes des observations se trouvent en table 52. Tous les ensembles définis possèdent un nombre d’observations suffisant pour être intégrés dans les analyses, donc aucun de ces groupes n’a été éliminé. Les fréquences ajustées par la méthode de l’expression count E.C. pour chaque variable sont données en table 53. DENTS PERMANENTES Maxillaire 1 Absence congénitale 2 Nombre de radicules 3 Racine appendiciforme 4 Nombre de racines des prémolaires 5 Nombre de racines des molaires 6 Racine de Carabelli 7 Racine paramolaire 8 Racine idiopathique 9 Degré de rotation 10 Convexité vestibulaire 11 Sillon traversant 12 Tubercule distal 13 Cingulum vestibulaire 14 Gouttière vestibulaire 15 Crêtes linguales marginales 16 Tubercule dentaire 17 Crêtes vestibulaires marginales 18 Crête mésiale 19 Crête accessoire distale 20 Cuspides accessoires 21 Odontome 22 Extension de l'émail 23 Parastyle 24 Trait de Carabelli 25 Métacone 26 Hypocone 27 Métaconule 28 Tubercule distal accessoire 29 Tubercule mésial paracone 30 Tubercule mésial accessoire 31 Protoconule 32 Tubercule lingual paracone 202 M3 M2 M1 P2 P1 C ● I2 I1 ● ● ● ● ● ● Mandibule ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● 33 Absence congénitale 34 Nombre de radicules 35 Racine appendiciforme 36 Nombre racines canines 37 Racine de Tomes 38 Nombre racines molaires 39 Racine de Citroen 40 Racine paramolaire 41 Racine entomolaire 42 Racine idiopathique 43 Crêtes linguales marginales 44 Crêtes vestibulaires marginales 45 Crête accessoire distale 46 Odontome 47 Variation des cuspides linguales 48 Pli cuspide mésiolinguale 49 Fosse précuspidale 50 Crête trigonide intermédiaire 51 Crête trigonide distale 52 Nombre de cuspides 53 Dessin du sillon 54 Protostylide 55-56 Hypoconulide-Entoconulide 57 Métaconulide 58 Tubercule paracone 59 Tubercule de Citroen M3 M2 M1 P2 P1 C I2 I1 ● ● ● ● ● ● ● ● ● ● ● ● ● Figure 96 : Représentation synthétique des dents clés retenues pour les analyses sexes séparés (n=44). L’analyse des données 7.2.5.3. Les variables mobilisées dans les analyses Pour cette partie de l’étude, 44 variables sur les 79 traits localisés sur les dents clés ont été retenues. 31 d’entre elles s’observent sur le maxillaire et 13 sur la mandibule (figure 96). Les fréquences E.C. sont données table 54. Les traits localisés sur les couronnes ainsi que sur les canines supérieures et les molaires sont, en général, mieux représentés. Les profils d’expression des variables observées sur les dents clés montrent des tendances similaires (figure 97). Il est tout de même possible de distinguer des divergences entre les groupes. Les profils des hommes cordés et campaniformes montrent l’allure la plus proche. Les deux ensembles unetices - hommes et femmes - présentent également des profils similaires. Ils se distinguent par la fréquence peu élevée de deux traits (métacone - T25 - et hypocone - T26 -) par rapport à celle rencontrée pour les quatre autres groupes cordés et campaniformes. Des valeurs plus élevées pour trois variables (Carabelli - T24 -, tubercule accessoire distal - T28 -, crêtes vestibulaires marginales - T43 -) peuvent être constatées pour les femmes cordées et campaniformes. 0.70 T25M1 T20P1 T2I1 T20P1 T2I1 0.00 T23M1 T23M1 0.00 T26M1 0.10 0.20 0.30 0.40 Cordé-homme Cordé homme 0.50 0.60 0.712 0.70 T17P1 T57M1 T18C T17I1 T17I1 T17P1 T22P1 T18C T57M1 T28M1 T22P1 T27M1 T27M1 T28M1 T29M1 T29M1 T1I2 T32M1 T32M1 T17C T58M1 T58M1 T17C T59M1 T59M1 T1I2 T11I1 T31M1 T8M1 T45C T31M1 T45C T11I1 T22M1 T22M1 T8M1 T1M3 T30M1 T24M1 T24M1 T30M1 T54M1 T54M1 T1M3 T19C T16I1 T19C T16I1 T10I1 T15C T16C T10I1 T16C T33M3 T13I1 T13I1 T15C T43I2 T43I2 T33M3 T4P1 T53M1 T53M1 T47P2 T47P2 T4P1 T2P1 T52M1 T34M1 T2M1 T52M1 T2M1 T2P1 T55-56M1 T55-56M1 T34M1 T5M1 T38M1 T5M1 T38M1 0.10 0.20 0.30 0.40 Cordé-femme Cordé femme 0.50 0.60 0.840 0.853 T26M1 T25M1 Profils d’expression des différents groupes définis pour l’analyse sexes séparés Figure 97 : Profil de chaque groupe testé dans les analyses, élaboré à partir des expression count des dents clés retenues pour l’étude sexes séparés (n=44). Liste des variables selon l’ordre dans le profil, de haut en bas: T26M1-T25M1-T5M1-T38M1-T55-56M1-T2M1-T52M1-T2P1-T34M1-T47P2-T4P1-T53M1-T43I2-T13I1-T16CT10I1-T15C-T33M3-T19C-T16I1-T54M1-T24M1-T1M3-T30M1-T22M1-T45C-T8M1-T11I1-T31M1-T27M1T29M1-T32M1-T58M1-T59M1-T1I2-T17C-T28M1-T17P1-T17I1-T18C-T22P1-T2I1-T20P1-T23M1-T57M1 203 204 0.60 0.10 0.20 0.30 0.40 0.50 0.60 T15C T33M3 T19C T4P1 T53M1 T43I2 T13I1 T16C T10I1 T15C T33M3 T4P1 T53M1 T43I2 T13I1 T16C T10I1 T15C T33M3 0.70 T10I1 T47P2 T47P2 T31M1 T11I1 T54M1 T24M1 T1M3 T30M1 T22M1 T45C T31M1 T54M1 T24M1 T1M3 T30M1 T22M1 T45C T31M1 T17I1 T17I1 0.00 0.00 T2I1 T18C T18C T2I1 T22P1 T22P1 T20P1 T28M1 T28M1 T23M1 T17C T17C T20P1 T1I2 T1I2 T23M1 T17P1 T27M1 T27M1 T17P1 T29M1 T29M1 T57M1 T32M1 T32M1 T17P1 T58M1 T58M1 T57M1 T17I1 T59M1 T59M1 0.00 T2I1 T20P1 T23M1 T57M1 T18C T22P1 T28M1 T17C T1I2 T27M1 T29M1 T32M1 T58M1 T59M1 T8M1 T11I1 T8M1 T11I1 T8M1 T45C T22M1 T30M1 T1M3 T24M1 T54M1 T16I1 T19C T16I1 T19C T16I1 T16C T13I1 T43I2 T53M1 T4P1 T47P2 T34M1 T2P1 T34M1 T2P1 T2M1 T55-56M1 T34M1 0.70 T52M1 T52M1 0.50 T2P1 T2M1 T2M1 0.40 T52M1 T55-56M1 T55-56M1 T38M1 T38M1 T38M1 T25M1 0.879 T5M1 T26M1 0.773 T5M1 0.30 Campa-femme Campaniforme femme T5M1 0.20 T25M1 0.781 T25M1 0.10 T26M1 0.791 T26M1 Campa-homme Campaniforme homme 0.10 0.20 0.30 0.40 0.50 Unet-homme Unetice homme 0.60 0.70 0.00 T2I1 T20P1 T23M1 T57M1 T17P1 T17I1 T18C T22P1 T28M1 T17C T1I2 T27M1 T29M1 T32M1 T58M1 T59M1 T8M1 T11I1 T31M1 T45C T22M1 T30M1 T1M3 T24M1 T54M1 T16I1 T19C T33M3 T15C T10I1 T16C T13I1 T43I2 T53M1 T4P1 T47P2 T34M1 T2P1 T52M1 T2M1 T55-56M1 T38M1 T5M1 T25M1 T26M1 0.10 0.20 0.30 0.40 0.50 Unet-femme Unetice femme 0.60 0.70 Chapitre 7 L’analyse des données 7.2.5.4. Les analyses Les premières analyses concernent uniquement les individus sexés pour les trois périodes étudiées (Cordé, Campaniforme et Unetice). -0.5 95 Les résultats des analyses multivariées sont des plus intéressants et présentent des configurations analogues (figure 98). L’analyse ascendante hiérarchique complétée par un bootstrap présente deux regroupements robustes (figure 98a). 100 84 4 90 80 Le premier est constitué des hommes et des femmes unetices. Le second réunissant les ensembles Cordé et Campaniforme. Au sein de cette configuration, les hommes et les femmes de chaque culture se rassemblent deux à deux. Le nuage de points issu du multidimensional scaling complété par un minimal spanning tree révèle une même tendance (figure 98b). L’unification des hommes et femmes datant du Bronze ancien est évidente. Les hommes cordés et campaniformes sont à nouveau très proches. Les femmes, quant à elles, sont reliées aux hommes campaniformes et semblent s’isoler d’un groupe à l’autre. L’analyse en composantes principales reproduit les éléments mis en évidence par les autres analyses. En effet, le nuage de points présente le regroupement des ensembles d’Unetice, la proximité des hommes cordés et campaniformes, et l’éclatement de la position des femmes de ces deux cultures (figure 98c). -a- 0 0 ■ Unet-homme ■ Unet-femme ● Cordé-homme ▼ Campa-homme Cordé-femme Campa-femme -bCampa-femme 0.2 Unet-femme ■ ▼Campa-homme 0 ■ Unet-homme ● Cordé-homme Cordé-femme -0.2 -0.48 0 0.48 -cCordé-femme 0.16 Que dire de ces résultats ? Les hommes des trois cultures sont proches et les femmes ont tendance à être plus hétérogènes. Les résultats de cette analyse sexes séparés sont compatibles avec ceux tirés de l’étude régionale. L’unité unetice est à nouveau mise en avant. Une plus grande variabilité des ensembles cordés et campaniformes se retrouve avec la position des Cordés (ici les hommes) que l’on pourrait considérer comme dénominateur commun à l’ensemble des populations. Cordé-homme ● Unet-homme ■ 0 ▼ ■ Campa-homme Unet-femme Campa-femme -0.2 -0.4 0 Figure 98 : Résultats des multivariées sexes séparés . 0.24 analyses a) Analyse ascendante hiérarchique, selon la méthode de Ward, complétée par un bootstrap . b) Multidimensional scaling complété par un minimal spanning tree (stress=0.00). c) Analyse en composantes principales (axe 1=71.8% et axe 2=11.3%). 205 Chapitre 7 Les deuxièmes analyses incluent trois groupes supplémentaires, soit les indéterminés de chaque culture. L’objectif étant de préciser la position des différents ensembles constatée lors de la première analyse. Les résultats des analyses multivariées présentent des tendances similaires (figure 99). -0.5 Campa-femme 87 ▼ 54 Campa-homme ▼ Campa-indéter 100 5 77 82 75 ■ Unet-homme ■ Unet-femme ■ Unet-indéter Cordé-indéter -bUnet-femme ■ 0.2 Unet-homme ■ Campa-femme Unet-indéter 0 Campa-homme▼ ■ ● Cordé-homme ▼ Campa-indéter Nous retrouvons les mêmes dispositions dans la représentation issue du multidimensional scaling complété par un minimal spanning tree (figure 99b). Cordé-femme Cordé-indéter -0.2 -0.4 0 0.32 -c- Cordé-indéter 0.16 Les sujets indéterminés cordés s’isolent, tout en étant reliés au groupe non sexé du Bronze ancien. Unet-indéter L’analyse en composantes principales indique à nouveau les éléments constatés lors des autres analyses. En effet, le nuage de points présente le regroupement des ensembles unetices, la position plus isolée des individus non sexés cordés et la localisation des hommes cordés en intermédiaire entre les deux autres cultures (figure 99c). ● Cordé-homme 76 Le premier comporte tous les ensembles du Bronze ancien (hommes, femmes et indéterminés). Les Cordés indéterminés se joignent à eux en formant un sous-groupe avec les sujets non sexés unetices. L’homogénéité du Bronze ancien est à nouveau mise en évidence. Les hommes cordés sont toujours en position intermédiaire entre Campaniforme et Unetice. Les sujets indéterminés campaniformes s’unissent aux hommes de la même période. Les femmes cordées et campaniformes présentent encore une certaine variabilité. Cordé-femme 76 L’analyse ascendante hiérarchique complétée par un bootstrap propose deux regroupements forts (figure 99a). Le second renferme les hommes et les femmes cordés et les trois ensembles campaniformes (hommes, femmes et indéterminés). A l’intérieur, on perçoit l’association des femmes des deux cultures. Les sujets indéterminés campaniformes se rattachent aux hommes de la même culture. Le groupe masculin cordé se joint à ce regroupement. -a- 0 0 ■ 0 Campa Cordé indéter femme ▼ Cordé-homme ● Campa ▼ Campa-homme femme Unet-homme ■ Unet-femme ■ -0.2 -0.4 0 0.24 Que dire de ces résultats ? Les mêmes éléments discutés lors de la première analyse sexes séparés sont à nouveau adéquats ici. L’intégration des sujets indéterminés des différentes cultures n’a pas amené de nouvelles informations. Néanmoins la position des individus non sexés cordés qui sont proches des ensembles unetices confirme l’implication du Cordé dans la mise en place du Bronze ancien. 206 Figure 99 : Résultats des analyses multivariées sexes séparés avec intégration des groupes indéterminés. a) Analyse ascendante hiérarchique, selon la méthode de Ward, complétée par un bootstrap . b) Multidimensional scaling complété par un minimal spanning tree (stress=0.090). c) Analyse en composantes principales (axe 1=51.5% et axe 2=29.2%). L’analyse des données 7.3. L’essentiel en bref Ce chapitre concernant l’analyse des données a comporté deux volets. Le premier a touché la présentation de la procédure suivie du traitement statistique préliminaires des données, le second s’est intéressé aux analyses proprement dites. • La procédure 1) Nous avons choisi de travailler avec des fréquences populationnelles. Nous avons donc calculé les incidences pour chaque variable au sein de chaque ensemble testé. Nous avons affaire à des variables dichotomiques et graduées, c’est pourquoi nous avons choisi de retenir la méthode dite de l’expression count mise en place par Turner (1985) qui permet d’obtenir une fréquence ajustée gardant l’ensemble de l’information fournie par les gradations. Elle s’effectue en deux temps : l’obtention d’une fréquence unique, puis l’introduction d’un facteur de correction prenant un compte la taille de l’échantillon. 2) Nous avons ensuite sélectionné les variables mobilisables dans les analyses. Lors du traitement préliminaire des données, nous avons vu que les traits s’exprimant au sein d’une classe dentaire ne sont pas indépendants les uns des autres. Nous avons donc retenu une dent par district dentaire que l’on appelle dent clé pour toutes les variables, y compris pour les traits qui ne semblent pas suivre le gradient d’expression (T15, T53 et T57). Le tubercule dentaire (T16) présente une interdépendance entre les classes dentaires, c’est pourquoi nous avons sélectionné une seule dent sur laquelle le trait s’exprime, généralement la canine. A l’issue de ce traitement statistique préliminaire des données, 79 variables sont intégrées aux analyses. 3) Nous avons retenu plusieurs types d’analyses multivariées : • L’analyse ascendante hiérarchique, selon l’algorithme de Ward, complétée par un bootstrap. L’analyse des proximités ou multidimensional scaling complété par un minimal spanning tree. L’analyse en composantes principales L’analyse des ensembles Plusieurs aspects ont été traités. Nous avons réalisé, pour toutes les aires étudiées (orientale, méridionale et à l’articulation - la Suisse -), une analyse individuelle de chaque région, puis une analyse du domaine en question. Nous avons comparé la Suisse une première fois au domaine oriental et une seconde au domaine méridional. Nous avons ensuite poursuivi les analyses par une étude comprenant uniquement les échantillons campaniformes de cinq régions traitées. Nous avons finalement clos la phase analytique par une étude du corpus tchèque sexes séparés. Quels sont les éléments à retenir ? Le domaine oriental Les analyses pour la Bohême se basent sur 13 ensembles (4 cordés, 4 campaniformes et 5 unetices) et 54 variables. Les différentes analyses défendent l’idée de populations plus hétérogènes lors de l’Enéolithique final en Bohême qui traduirait probablement une mobilité ou des échanges populationnels plus accrus pendant ces périodes. Le début du Bronze ancien, en revanche, révèle une forte cohésion des groupes étudiés présupposant un certain hermétisme aux influences externes. La position intermédiaire du Cordé plaiderait en faveur de son implication dans la mise en place du Campaniforme et de la culture d’Unetice. Les analyses pour la Hongrie sont réalisées à partir de 4 ensembles (Campaniforme, Obedà-Pitvaros, Perjamos et Nagyrev) et 35 variables. La culture Nagyrev semble avoir une position forte et centrale 207 Chapitre 7 entre les différentes cultures étudiées. Le Campaniforme s'individualise sur l’ensemble des représentations. Il ne s’avère proche ni de la culture méridionale qui lui est contemporaine (ObedàPitvaros), ni par le Nagyrev qui lui est en partie contemporain - dans sa phase ancienne - et qui lui succède. Les analyses du domaine oriental se basent sur 17 ensembles (pour la Bohême les 4 cordés, 4 campaniformes et 5 unetices, pour la Hongrie 4 groupes) et 30 variables. Pour la Bohême, deux regroupements (Unetice/Cordé oriental et Cordé/Campaniforme) se distinguent. La position des groupes hongrois au sein de cette configuration est très intéressante puisque le Campaniforme se rattacherait modérément au premier groupe et les trois autres ensembles - Nagyrev, Obedà-Pitvaros et Perjamos - au second groupe. Le domaine méridional Les analyses pour le sud de la France se basent sur 7 ensembles (2 Néolithique final, 2 Chalcolithique, 1 Campaniforme, 1 Chalcolithique/Campaniforme et 1 Chalcolithique/Bronze ancien) et 57 variables. Les gisements du Néolithique final et du Chalcolithique présentent une situation plus hétérogène. En revanche, les gisements récents s’isolent. Les analyses pour le nord de l’Espagne sont réalisées à partir de 7 ensembles (3 Néolithique final, 2 Chalcolithique, 1 Campaniforme et 1 Néolithique final/Campaniforme) et 17 variables. Les différents échantillons appartenant au Néolithique final paraissent homogènes. Les ensembles récents, en revanche, semblent montrer une plus grande variabilité. La position des Campaniformes en intermédiaire entre les populations du Néolithique final et du Chalcolithique se révèle fort intéressante. Les analyses du domaine méridional ont été effectuées à partir de 13 ensembles (7 français et 6 espagnols) et 30 variables. La situation mise en évidence dans ce domaine est intéressante puisqu’on retrouve des différences plus marquées pour les périodes antérieures et une certaine cohésion des populations les plus récentes traduisant probablement des échanges plus marqués entre elles. Le Campaniforme au sein de cette configuration se situe entre ces deux courants. A l’articulation entre les deux domaines Les analyses pour la Suisse se basent sur 10 ensembles (5 groupes Néolithique moyen, 2 Néolithique récent, 2 Campaniforme et 1 Bronze ancien) et 49 variables. Les différentes analyses s’accordent pour défendre l’idée d’une certaine harmonie des populations du Néolithique moyen et d’une variabilité des ensembles tardifs qui traduirait probablement une mobilité ou un apport populationnel modéré, et ce, dès le Néolithique récent. Les analyses des relations de la Suisse avec le domaine oriental sont réalisées à partir de 22 ensembles (5 groupes suisses, 13 tchèques et 4 hongrois) et 30 variables. Nous constatons les mêmes tendances distinguées lors des analyses régionales. Les sites suisses ne se mêlent pas au domaine oriental et les Campaniformes hongrois semblent bien plus proches de leurs contemporains helvétiques. Les analyses des relations de la Suisse avec le domaine méridional sont effectuées à partir de 18 ensembles (5 groupes suisses, 7 français et 6 espagnols) et 30 variables. Les échantillons helvétiques se regroupent fortement sur l’ensemble des configurations. Ils se rattachent aux périodes récentes ibériques et françaises. Si les groupes anciens du domaine méridional présentent une plus grande variabilité, traduisant probablement une mobilité ou des échanges populationnels moindres pendant ces périodes, ceci ne semble pas être le cas en Suisse et pour les ensembles tardifs méridionaux où l’homogénéité apparente permettrait d’avancer une accentuation des influences externes. 208 L’analyse des données Les relations entre les populations campaniformes Cette analyse se base sur 11 ensembles (4 groupes tchèques, 1 hongrois, 2 suisses, 2 français et 2 espagnols) et 30 variables. Une dichotomie certaine et franche des populations campaniformes - domaine oriental et domaine méridional - se révèle au sein des analyses. Les Campaniformes helvétiques se rattachent clairement au domaine méridional. Les échanges et les influences externes semblent majeurs dans l’aire occidentale campaniforme. La Bohême : une analyse sexes séparés Cette dernière analyse comprend 6 groupes (hommes et femmes cordés, campaniformes et unetices) et 44 variables. Les hommes des trois cultures sont proches et les femmes ont tendance à être plus hétérogènes. Les résultats sont assimilables à ceux de l’étude régionale : une unité unetice, une plus grande variabilité des ensembles cordés et campaniformes et une position des Cordés - les hommes - en dénominateur commun fédérant l’ensemble des populations. 209 Chapitre 8 De la discussion à l’interprétation Nous avons étudié cinq régions à occupation campaniforme, cinq instantanés différents provenant des aires orientale (Bohême et Hongrie) et méridionale (sud de la France et nord de l’Espagne) du phénomène, ainsi que de Suisse, en position médiane entre ces deux grands courants. Afin d’aborder la dynamique des peuplements de l’Europe au 3e millénaire avant notre ère, il est indispensable de procéder graduellement à l’interprétation des résultats. Dans un premier temps, l’échelle locale est retenue pour comprendre, d’un point de vue chronologique, les relations entre les populations. Dans un deuxième temps, les résultats fournis par les études interrégionales sont mobilisés afin de saisir les connexions spatio-temporelles entre les différentes cultures. Finalement, les analyses ponctuelles entre échantillons campaniformes ou sexes séparés - permettent de proposer un modèle de diffusion du complexe campaniforme. A l’échelle de chaque région, nous devons clarifier un certain nombre de points : Dans un premier temps, il est nécessaire d’établir : • • (1) les relations entre les ensembles précampaniformes (2) les liens entre les échantillons campaniformes (3) afin d’évaluer le rôle du fonds précampaniforme dans l’émergence du Campaniforme. (4) Ensuite, nous mesurons le fonds campaniforme dans la mise en place du Bronze ancien. (5) Ces différents éléments doivent nous conduire, pour chaque région, à déterminer s’il y a eu, lors du 3e millénaire avant notre ère, un renouvellement de la population et à quel moment il est possible de le mettre en évidence. Selon les caractéristiques de chaque région, nous pouvons répondre intégralement ou partiellement à ces questions. A une échelle interrégionale, nous devons préciser : • • • • (6) les rapports entre les populations appartenant à un même domaine (7) les relations du territoire helvétique avec les aires orientale et méridionale (8) les connexions entre les différents échantillons campaniformes (9) les comportements sexes différenciés de l’échantillon tchèque Ces éléments doivent permettre de proposer une dynamique des peuplements à l’échelle du phénomène. 8.1. Une discussion à l’échelle régionale 8.1.1. La Bohême Nous avons inclus dans les analyses pour la Bohême, le Campaniforme, son substrat, - en partie contemporain -, le Cordé, et la période qui lui succède, la culture d’Unetice. Les résultats des différentes analyses révèlent une situation relativement nette (figure 68). Ainsi, pour la morphologie dentaire : (1) Les groupes précampaniformes, les Cordés, forment un ensemble cohérent. Par rapport à l’unité du Bronze ancien, ils paraissent néanmoins moins homogènes. (2) Les Campaniformes présentent une variabilité plus importante. L’ensemble occidental (BWca) se distingue de ses congénères, tout en restant proche des Cordés. (3) Le Cordé est - sans aucun doute - impliqué dans la mise en place du Campaniforme. 211 Chapitre 8 (4) Le Campaniforme, quant à lui, ne semble pas engagé dans l’émergence de la culture d’Unetice. En effet, ces deux cultures se distinguent clairement sur toutes les configurations. (5) Les données fournies par l’anthropologie dentaire ne plaident, à aucun moment, en faveur d’un renouvellement de la population. Néanmoins, il est possible de mettre en évidence une certaine mobilité ou des échanges populationnels pendant le Cordé. Ce phénomène semble se renforcer pendant le Campaniforme. Ceci ne se vérifie pas pour la culture d’Unetice puisqu’elle dévoile une cohésion importante indépendamment de la localisation géographique des différents ensembles. Si le Campaniforme et le Bronze ancien évoluent indépendamment l’un de l’autre, l’implication du fonds cordé apparaît bel et bien chez chacun d’entre eux. Les autres composantes culturelles ou biologiques livrent en partie des résultats similaires. Les différentes données (céramique, rituel funéraire, structures domestiques...) s’accordent pour dire que le Cordé contribue fortement à la mise en place du Campaniforme en Bohême. Si la céramique décorée campaniforme évoque celle du reste du domaine, la céramique commune est de tradition régionale. Les types sont empruntés au fonds local cordé et à diverses cultures d’Europe centrale (Besse 2001, 2003). On pense qu’une évolution interne de la société du Cordé à conduit au Campaniforme (Turek et Peska 2001). Les rituels funéraires similaires, suivant chacun leurs propres règles, appuient cette hypothèse. De plus, les Cordés et les Campaniformes côtoient parfois la même nécropole, voire la même la tombe. Les structures domestiques, même si elles sont rares - tant pour le Cordé que le Campaniforme -, ne se distinguent pas de manière significative (Turek et Peska 2001). On ne peut donc nier cette proximité. Par ailleurs, l’analyse des pâtes des céramiques campaniformes montre que les poteries sont produites localement et que parfois des déplacements de faible ampleur peuvent être mis en évidence (Rehman et al. 1992). Ces éléments n’excluent pas un apport populationnel pendant le Campaniforme, lequel, selon les données dentaires est présent, et ce dès le Cordé (quoique de manière plus modérée). Cet apport externe a été mesuré lors des analyses entre échantillons campaniformes et sexes séparés qui sont discutés au chapitre 8.2. Les données anthropologiques se sont souvent basées sur des échantillons constitués d’après le sexe des individus (Czarnetzki 1984, Price et al. 2004) et leurs résultats montrent une certaine mobilité des individus campaniformes qui diffère selon l’approche utilisée. Nous y reviendrons. Il n’est pas fréquent de rencontrer dans la littérature des comparaisons entre les cultures Cordé et Unetice. On admet une continuité entre le Cordé et le Campaniforme (Turek et Peska 2001, Vander Linden 2006...), et ce dernier avec la culture d’Unetice (Zápotock 1994, Krutova 2003...). De ce fait, le Campaniforme est confronté soit à l’une soit à l’autre. Et l’exercice se révèle d’autant plus complexe puisqu’on recherche avant tout ce qui les fédère. Nous avons une continuité du système symbolique du Cordé au Bronze ancien, néanmoins il est possible de relever des spécificités selon les cultures et les régions (Krutova 2003). Certains chercheurs pensent qu’il existe un lien étroit entre le Campaniforme et le proto-Unetice (Hrala et al. 1994). De plus, il ne semble pas exclu qu’il puisse y avoir eu une coexistence du Cordé, du Campaniforme et du protoUnetice. En revanche, lors du développement de la culture d’Unetice, elle est seule dans l’aire étudiée (Hrala et al. 1994). Nous cherchons à comprendre le développement autonome, d’après la morphologie dentaire, des cultures Campaniforme et Unetice. Les décors et les formes des céramiques décorées campaniformes n’ont aucun impact sur la culture d’Unetice, en revanche la céramique commune les relie (Turek et Peska 2001). Cette céramique est déjà de tradition cordée pour le Campaniforme. On peut également évoquer la présence, en contexte funéraire, des haches-marteaux que l’on retrouve uniquement au Cordé et au Bronze ancien et le mobilier d’archer que l’on retrouve exclusivement au Campaniforme (Krutova 2003). Ainsi, les différentes composantes concordent en faveur d’une évolution au sein d’une même société lors du passage du Cordé au Campaniforme. En revanche, le fonds local sur lequel se met en place la culture d’Unetice serait probablement à rechercher dans le Cordé et non dans le Campaniforme, issu lui-même du Cordé ! 212 De la discussion à l’interprétation 8.1.2. La Hongrie L’interprétation des résultats pour la Hongrie est moins précise puisque nous avons travaillé à partir des fréquences par cultures. De plus, le substrat sur lequel le Campaniforme se développe - la culture de Makó - n’est pas inclus dans les analyses. Ainsi, cette région est abordée avec des cultures qui sont soit contemporaines (Obedà-Pitvaros), soit successives (Perjamos et Nagyrev) au Campaniforme. Les résultats des différentes analyses ne permettent d’aborder que partiellement les éléments à clarifier, néanmoins ils fournissent des éléments intéressants (figure 72) : (1) Les relations entre les groupes précampaniformes n’ont pas pu faire l’objet d’une évaluation. (2) Les échantillons campaniformes sont regroupés en un seul ensemble, ainsi les comportements internes au Campaniforme ne sont pas identifiables. (3) Sur la base des point précédents, la morphologie dentaire n’est pas en mesure de définir si le fonds culturel local - le Mako - est à l’origine du Campaniforme. (4) Il est toutefois possible de déterminer les relations du Campaniforme avec les groupes culturels contemporains ou successifs. Il se distingue franchement non seulement de la culture contemporaine méridionale d’Obedà-Pitvaros, mais également des groupes postérieurs de Perjamos et de Nagyrev. La culture la moins éloignée du Campaniforme reste tout de même le Nagyrev. (5) Les données fournies par l’anthropologie dentaire fournissent une image intéressante des contacts entretenus par le Campaniforme avec les autres groupes culturels régionaux. Le Campaniforme se détache clairement sur l’ensemble des configurations. Le Nagyrev, en revanche, semble détenir une position forte et centrale des relations entre les différentes cultures étudiées. Même si nous n’avons pas pu travailler sur le rôle du fonds local dans la mise en place du Campaniforme hongrois, les différences relevées - qui sont tout de même relativement marquées - plaideraient en faveur d’un renouvellement de la population. Les autres composantes culturelles ou biologiques proposent des résultats plutôt mitigés. Nous avons vu que l’intérêt du Campaniforme de cette région est multiple. Il s’agit de l’expression la plus orientale du phénomène, le fonds sur lequel le Campaniforme s’installe n’est pas le Cordé, son extension se limite aux environs de Budapest et il se place dans une phase moyenne du Bronze ancien. L’incompréhension que suscite sa présence en territoire hongrois provoque de grandes discussions. Les différents éléments provenant des composantes archéologiques montrent à la fois de grandes affinités et des divergences importantes avec les cultures locales - notamment avec la culture de Nagyrev. En effet, le matériel strictement campaniforme se retrouve principalement en contexte funéraire. Les éléments métalliques, que l’on rencontre dans les gisements domestiques, ont des typologies proches de celles du reste du phénomène. Tous ces éléments sont fort similaires aux découvertes campaniformes d’Autriche et de Moravie (Kalicz-Schreiber et Kalicz 2001). Les autres composantes culturelles présentent des affinités avec les traditions du fonds local et du Nagyrev. L’analyse des pâtes montre que la céramique est produite sur place (Rehman et al. 1992). De même, la céramique commune et, en particulier, le pichet à anse est de tradition locale. Les éléments de la culture matérielle ne sont pas seuls à comporter des influences d’origines multiples. Les structures funéraires sont, non seulement, des incinérations de tradition locale, mais également des inhumations évoquant le reste du domaine campaniforme et considérées comme an alien tradition (Kalicz-Schreiber et Kalicz 2001, p. 442). Les données provenant de l’anthropologie biologique (biométrie et géochimie isotopique) convergent et attribuent généralement une origine extra locale aux individus campaniformes hongrois. Ainsi, en réponse aux interrogations de R. Kalicz-Schreiber et N. Kalicz (2001) were the Bell Beakers as social indicators of the early Bronze age in Budapest ? l’anthropologie dentaire défendrait de préférence l’idée d’une nouvelle population venue s’installer sur les bords du Danube, mais qui aurait eu des contacts et des échanges privilégiés avec les groupes locaux, notamment avec la culture de Nagyrev. 213 Chapitre 8 8.1.3. Le sud de la France Nous avons inclus dans l’analyse régionale du sud de la France des gisements se situant entre le Néolithique final et le Bronze ancien. Il s’agit de sépultures collectives pour lesquelles les attributions culturelles ne sont pas toujours individualisées, c’est pourquoi certains ensembles contiennent deux périodes distinctes. Les configurations proposées par les analyses révèlent une situation intéressante des relations entre les échantillons (figure 78) : (1) Les groupes précampaniformes, les échantillons Néolithique final/Chalcolithique, se concentrent sur les différentes représentations, où il est tout de même possible de mettre en évidence une certaine variabilité des ensembles. Cette situation est d’autant plus évidente pour le gisement de Roaix avec deux occupations précampaniformes distinctes. (2) Pour le Campaniforme, on peut mesurer une légère hétérogénéité des deux ensembles intégrés dans les analyses. (3) Le fonds néolithique final/chalcolithique local ne semble pas impliqué dans la mise en place du Campaniforme. (4) Les deux groupes campaniformes se rattachent au gisement de Cudières, contenant des mélanges Chalcolithique et Bronze ancien. Il est difficile, dans le contexte de l’analyse que nous avons menée, d’évaluer si le Campaniforme est impliqué dans la mise en place du Bronze ancien. Nous pouvons simplement mettre en évidence la proximité des ensembles récents du sud de la France. (5) Les données fournies par l’anthropologie dentaire s’orienteraient de préférence vers un certain renouvellement de la population avec l’émergence du Campaniforme. Les gisements précampaniformes présentent, à l’évidence, une situation hétérogène reflétant probablement la configuration pluriculturelle existante. En revanche, les gisements mixtes ultérieurs (Les Peyraoutes et Cudières) s’isolent en gardant une proximité avec l’ensemble campaniforme de Villedubert, ce dernier se positionnant en intermédiaire entre les regroupements antérieurs et récents. Les composantes culturelles permettent de préciser la situation. L’étude de la céramique commune met en évidence des morphologies nouvelles que l’on ne retrouve pas dans le fonds local (Besse 2001, 2003). L’approche technologique propose que l’on soit en présence de transferts techniques et d’un savoir-faire qui n’est pas à rechercher dans le substrat local (Convertini 1996). La périodisation de la céramique décorée en quatre stades permet de mettre en évidence une évolution du Campaniforme : une phase ancienne de rupture avec les traditions locales, une phase d’intégration et de cohabitation, une phase de différenciation et une dernière phase de distinction marquant la fin du complexe, voire le début du Bronze ancien pour certains (Lemercier 2002, 2004). L’analyse de l’industrie lithique livre des résultats similaires avec une bipartition du Campaniforme (une rupture suivie par une phase d’intégration et de diffusion) (Furestier 2005). Les structures funéraires sont variées. Plusieurs inhumations individuelles campaniformes sont mises en évidence, néanmoins le rituel le plus fréquent est le réemploi de sépultures collectives du Néolithique final (Besse et Desideri 2005). Les structures domestiques sont de deux types : les gisements « purs » strictement campaniformes et les habitats « mixtes » comportant des vestiges appartenant aux diverses traditions locales du Néolithique final et au Campaniforme (Besse et Desideri 2005). Les données biologiques, notamment les données métriques du crâne, ne fournissent actuellement pour cette région aucun élément permettant d’alimenter la recherche dans une perspective d’histoire du peuplement. Ainsi, les différentes composantes vont dans le sens d’un renouvellement de la population avec l’émergence du Campaniforme. Les données archéologiques permettent de préciser les modalités de son installation avec une phase marquée par l’apparition d’un mobilier nouveau, suivie par une phase de cohabitation plus ou moins soutenue avec les traditions locales du Néolithique final (Lemercier 2002, 2004, Furestier 2005). 214 De la discussion à l’interprétation 8.1.4. Le nord de l’Espagne Le nord de l’Espagne est abordé par diverses périodes précampaniformes (Néolithique final et Chalcolithique) et par le Campaniforme. L’âge du Bronze n’a pas été retenu dans les analyses pour des raisons d’effectifs insuffisants. Un gisement présente une situation mixte Néolithique final/Campaniforme. Les résultats des différentes analyses (figure 82) dévoilent une situation relativement claire. Ainsi, pour les données dentaires : (1) Les groupes précampaniformes présentent des comportements différents. Une grande cohésion des populations du Néolithique final est perceptible. Le Chalcolithique, au contraire, semble bien plus hétérogène. (2) Les deux ensembles campaniformes ne semblent pas proches. L’un des deux gisements contient un mélange d’occupations qui pourrait être à l’origine de cette divergence. (3) Le fonds local, composé par le Néolithique final et le Chalcolithique, est vraisemblablement impliqué dans l’émergence du Campaniforme. (4) Le fonds campaniforme n’a pas pu être évalué dans la mise en place du Bronze ancien puisque ce dernier n’a pas été intégré dans l’étude. (5) Les données fournies par l’anthropologie dentaire ne plaident pas en faveur d’une rupture du peuplement lors de l’émergence du Campaniforme. Une homogénéité importante au Néolithique final semble s’opposer à une variabilité certaine au Chalcolithique. La position des Campaniformes en intermédiaire entre les populations du Néolithique final et du Chalcolithique semble signifier que toutes deux ont été engagées dans la mise en place du Campaniforme. Les composantes culturelles et biologiques offrent des résultats similaires. Le fonds local semble avoir un rôle important dans la mise en place du Campaniforme. En effet, un fort impact du substrat dans la morphologie des différentes céramiques est perceptible. De plus, les analyses des pâtes montrent que la production est réalisée avec des matériaux régionaux (Millàn et Arribas 1994). La métallurgie est connue bien avant l’émergence du Campaniforme. De plus, elle est bien attestée pendant le Campaniforme et s’inscrit fortement dans le fonds néolithique récent local (Garrido-Pena et al. 2005). Les traces de bière mises en évidence dans les céramiques permettent de mettre à nouveau sur les rangs la relation entre céramique campaniforme et contenant rituel (Garrido-Pena 2005). Les structures et le matériel funéraires présentent une variabilité certaine (Harrison et Mederos Martin 2001). On note tout de même une persistance des traditions locales pendant le Campaniforme. Les structures funéraires, réutilisant des monuments du Néolithique final et du Chalcolithique, présentent un rituel standardisé des sépultures campaniformes témoignant, pour certains, d’un traitement particulier du défunt ayant une signification sociale (Garrido-Pena et al. 2005). L’étude des structures domestiques montre qu’aucune différence des systèmes économiques entre le fonds régional local précampaniforme et le Campaniforme ne peut être mise en évidence (Harrison et Mederos Martin 2001). Les analyses anthropologiques basées sur les données métriques crâniennes - relevant en général sur le territoire occupé par les Campaniformes une morphologie particulière de ces derniers - mettent en évidence une morphologie empruntée au fonds local méridional (Garralda 1979). Ainsi, une continuité de peuplement avec l’émergence du Campaniforme peut être mise en évidence sur l’ensemble des composantes étudiées. La situation intermédiaire du Campaniforme entre the Late Neolithic communal societies and the Bronze Age chiefdoms could also explain the, to a certain extend transitional, character of Beaker burial rites (Garrido-Pena 2005, p. 56). Pour certains, notamment R. Garrido-Pena, M. Rojo Guerra et I. Garcia Martinez de Lagran (2005), la céramique décorée campaniforme se révèle un bien socialement valorisé intégré dans des stratégies de légitimation de dirigeants des sociétés du Chalcolithique dans un contexte de transformations des processus socioéconomiques. 215 Chapitre 8 8.1.5. La Suisse Les échantillons mobilisés au sein de l’étude pour la Suisse se situent chronologiquement entre le Néolithique moyen et le Bronze ancien. Les configurations proposées par les analyses exposent une image claire des relations entre les différentes populations de la fin du Néolithique helvétique (figure 88). Ainsi, pour les données dentaires : (1) Les groupes précampaniformes dévoilent des comportements différents. Les populations du Néolithique moyen forment un noyau homogène. En revanche, les ensembles du Néolithique récent montrent une plus grande variabilité. (2) Pour le Campaniforme, une certaine hétérogénéité des deux ensembles intégrés dans les analyses peut être mise en évidence. (3) Le fonds néolithique récent est impliqué dans l’émergence du Campaniforme, mais il ne semble pas en être l’unique interprète. (4) Le Campaniforme, quant à lui, ne semble pas engagé dans la mise en place du Bronze ancien. En effet, ces deux cultures se distinguent sur toutes les configurations. (5) Les données fournies par l’anthropologie dentaire plaident en faveur d’un renouvellement partiel de la population. La variabilité des ensembles tardifs va dans le sens d’une mobilité ou d’un apport populationnel modéré pendant ces différentes périodes. Pour le Bronze ancien la situation n’est pas aussi claire. Il semble tout de même plus proche du fonds néolithique moyen que de celui des périodes récentes. La cohésion des populations du Néolithique moyen en territoire helvétique en revanche paraît certaine. Les composantes culturelles et biologiques précisent l’apport des influences externes. La céramique marque une rupture relativement nette, celle du Néolithique récent fait place aux gobelets décorés et à des formes nouvelles (Besse 2001, 2003). Les objets en cuivre sont très nombreux au Néolithique récent, tandis qu’au Campaniforme on ne dénombre qu’une seule alène (Cattin en cours). La différence dans ce cas est d’ordre quantitatif. Si le Néolithique récent est marqué par une forte occupation littorale dans la région des Trois-Lacs et autour du lac Léman, le début du Campaniforme, quant à lui, est marqué par l’abandon des sites lacustres. Il s’agit d’un phénomène observé dans tous les lacs suisses et du Jura français. Cet abandon ne semble pas à mettre en relation avec un changement climatique qui aurait alors forcé les Campaniformes à changer d’environnement pour s’y installer (M. Piguet, communication personnelle). En ce qui concerne les espaces funéraires, les Campaniformes continuent à utiliser ceux du Néolithique récent. L’exceptionnelle aire mégalithique du Petit-Chasseur à Sion en est un excellent exemple. Les occupants campaniformes édifient leurs propres monuments au sein de l’aire mégalithique déjà fréquentée par leurs prédécesseurs. Ils construisent un certain nombre de dolmens et de cistes puis réemploient une structure antérieure. Ces comportements composites résultent d’influences variées. On distingue des influences méditerranéennes notamment par les gobelets maritimes - sensu stricto -, les grandes lames en silex exogène et les réoccupations des dolmens. Des influences orientales sont mises en évidence par des décors et des formes de la céramique commune, par les pointes de flèche à base concave, par la présence de sépultures individuelles et par certaines parures en or et en argent (Besse 1998b). La délocalisation des habitats semble une volonté de démarcation que l’on repère dans le domaine oriental, attestant, par la suite, d’une originalité locale. Les études anthropologiques attribuent, en général, l’émergence du Campaniforme en territoire helvétique à une certaine mobilité des individus (Menk 1979, Chiaradia et al. 2003, Desideri et Eades 2004...). Ainsi, les différentes composantes indiquent que le Campaniforme ne témoigne pas d’une réelle rupture avec les périodes précédentes, mais plutôt d’une accentuation des changements liés aux influences extérieures. Ces influences sont la conséquence d’un certain apport qui est à la fois populationnel et idéologique. Le Campaniforme est donc un moment privilégié pour les contacts soutenus avec l’Europe, tant méridionale qu’orientale, ce qui génère, en Suisse occidentale, un groupe culturel composite. 216 De la discussion à l’interprétation 8.2. Une discussion à grande échelle Cette partie de la discussion contribue à mettre en évidence les relations entre les populations à différentes échelles spatio-temporelles. (6) Intéressons-nous dans un premier temps aux rapports entre les populations appartenant à un même domaine. Les résultats concernant les relations entre les échantillons provenant du domaine oriental offrent une configuration intéressante, néanmoins inattendue (figure 74). Les comportements des différents ensembles culturels sont à nouveau identifiés (rôle central du Cordé et développement indépendant du Campaniforme et de la culture d’Unetice). Si les Hongrois non campaniformes s’intègrent parmi leurs congénères tchèques, le Campaniforme se distingue clairement. Ainsi, ni le fonds local hongrois, ni le fonds régional oriental ne paraissent impliqués dans son émergence. Ceci va à l’encontre des hypothèses émises par un certain nombre de chercheurs qui attribue à l’enclave du Campaniforme-Csepel une origine en Bohême, ou du moins provenant de Tchéquie (Endrödi 2003). Les résultats concernant les liens entre les populations provenant du domaine méridional révèlent une situation analogue à celle rencontrée lors des analyses régionales (figure 84). De plus, ils fournissent des éléments de compréhension importants. L’hétérogénéité des ensembles du Néolithique final et du Chalcolithique pourrait être mise en parallèle avec la diversité culturelle présente notamment sur l’ensemble du Midi de la France, région que l’on compare souvent à une aire de compétition sociale à la fin du Néolithique (Vander Linden 2006). Le regroupement des populations tardives - Campaniforme et Bronze ancien - sur l’ensemble du domaine, combiné aux résultats des analyses régionales, permettent de préciser les interprétations concernant les modalités de l’émergence du Campaniforme dans ces deux régions. Le Campaniforme de la Meseta possède un fort ancrage local. Le sud de la France se distingue de son fonds régional et se rapproche clairement des populations contemporaines ibériques. Ces éléments vont dans le sens des hypothèses émises par certains chercheurs, en particulier O. Lemercier (2002, 2004), pensant qu’un apport de nouvelles populations venant de péninsule Ibérique pourrait être à l’origine du Campaniforme dans cette région. De plus, si seule l’analyse locale du sud de la France ne permet pas de déterminer l’influence du Campaniforme dans la mise en place du Bronze ancien, à l’échelle du domaine, son implication semble effective. (7) Abordons ensuite les relations du territoire helvétique avec les aires orientale et méridionale. La position de la Suisse entre les deux domaines fournit des éléments de compréhension nouveaux. Les sites helvétiques ne se mélangent pas au domaine oriental (figure 90), en revanche ils s’intègrent en parfaite harmonie au sein du domaine méridional (figure 92). L’analyse régionale de la Suisse a montré qu’un apport populationnel externe modéré est présent dès le Néolithique récent. L’axe des influences extérieures est méridional, allant à l'opposé des interprétations issues non seulement des données anthropologiques anciennes (Menk 1979, 1981), mais également des scénarii du peuplement campaniforme dans cette région annonçant des influences idéologiques méridionales et des mouvements de populations orientaux (Gallay 2006). La Suisse est un territoire présentant la dichotomie des traditions ouest-est. Et l’échantillon sélectionné pour les analyses est constitué principalement de gisements appartenant à la partie occidentale helvétique. Il n’est donc pas exclu qu’un apport populationnel provenant d’Europe centrale en Suisse soit présent, mais il se limiterait à l’aire orientale de la région. (8) Etudions également les connexions entre les Campaniformes européens. Les résultats des analyses qui ont concerné uniquement les échantillons campaniformes sont des plus parlants (figure 95). Il est possible de mettre en évidence deux centres populationnels correspondant aux deux domaines étudiés. Les ensembles appartenant au domaine méridional montrent une grande cohésion allant dans le sens d’influences et d’échanges plus importants au sein de la communauté campaniforme. Des mouvements qui pourraient correspondre aux déplacements de groupes d’individus depuis la péninsule Ibérique, puisque dans cette région aucun renouvellement de la population n’est décelable. La Suisse se rattache sans équivoque à ce domaine. 217 Chapitre 8 La situation dans l’aire orientale est différente. Les Campaniformes tchèques sont également regroupés mais de façon moins homogène. De ce fait, les modalités de diffusion ne semblent pas comparables au sein des deux domaines. Il n’est donc pas envisageable d’imaginer des mouvements liés à des groupes d’individus dans l’aire orientale. La situation du Campaniforme hongrois se précise, puisque les individus se situent en intermédiaire entre les deux domaines et semblent se rattacher au domaine méridional. (9) Enfin, testons le comportement sexes différenciés de l’échantillon tchèque. Cette analyse permet de préciser les modalités de diffusion du Campaniforme, mais uniquement au niveau de la région pour laquelle l’étude peut être réalisée, soit la Bohême. Dans l’aire orientale, le sexe d’un individu peut être attribué en fonction des règles d’inhumation (orientation des corps et matériel associé). Néanmoins, pour cette partie de l’étude, nous avons préféré les déterminations sexuelles fournies par les données anthropologiques. Les résultats des comportements sexes séparés sont comparables à ceux de l’étude régionale de la Bohême (figures 98 et 99). L'harmonie unetice et la variabilité des ensembles cordés et campaniformes sont à nouveau perceptibles. Néanmoins, la position forte et centrale de la culture Cordé, et en particulier des hommes, semble confirmer son statut de dénominateur commun et son implication dans la mise en place des périodes successives. De plus, l’homogénéité des hommes des trois cultures semble témoigner d’une stabilité masculine pendant l’Enéolithique en Bohême. Les femmes, en revanche, dévoilent une plus grande hétérogénéité. La proximité des hommes et des femmes unetices renforce l’idée d’une société du Bronze ancien plus fermée aux échanges ou aux apports populationnels, ce qui ne semble pas le cas ni pour le Cordé, ni pour le Campaniforme. Nos résultats vont à l’encontre des interprétations d’A. Czarnetzki (1984) établies à partir des données non métriques crâniennes de ces même populations, mettant en évidence des mouvements migratoires des hommes uniquement pendant le Campaniforme. En revanche les femmes montreraient une morphologie similaire à celles des femmes des deux autres périodes, allant dans le sens d’une proximité et d’un équilibre des individus de sexe féminin dans le temps. Nos interprétations sont, en revanche, proches de celles révélées par les analyses isotopiques du plomb et du strontium réalisées par D. Price et al. (2004). Leurs résultats révèlent une certaine mobilité pendant le Campaniforme sur l’ensemble de l’aire étudiée soit l’Europe centrale - et les femmes paraîtraient légèrement plus mobiles. La mobilité apparente des femmes pendant le Cordé et le Campaniforme permet d'approcher la question des réseaux d’alliances matrimoniales lors de l’Enéolithique dans l’aire orientale. L’étude de N. Brodie (2001) a déjà abordé cette problématique. En effet, ce chercheur a travaillé sur la relation entre la production de la céramique et la femme dans une perspective de compréhension du Campaniforme dans le nord et l’est de l’Europe. Il a mis en place un modèle d’identité féminine campaniforme en se basant sur les données ethnologiques issues des sociétés sédentaires et semi sédentaires du Pacifique lors des premiers contacts européens. Selon son modèle, l’Europe néolithique (occidentale) est en contact avec l’Europe chalcolithique (orientale). La volonté des populations de l’ouest d’acquérir non seulement les objets, mais également le savoir-faire métallurgique, a eu pour conséquence de modifier les stratégies de mariage. Ainsi, les femmes chalcolithiques orientales auraient intégré peu à peu l’ouest, et auraient de ce fait déplacé graduellement la frontière chalcolithique. Certains chercheurs, notamment R. Garrido-Pena (2005), pensent que si ce modèle est valable, il ne peut s’appliquer qu’à l’est et au nord du phénomène Campaniforme puisque la métallurgie est attestée bien avant le Campaniforme en péninsule Ibérique notamment. S’il ne paraît pas envisageable d’étendre ce modèle à l’ensemble du territoire campaniforme, les traits non métriques dentaires dévoilent des divergences importantes des femmes - cordées et campaniformes - par rapport à l’homogénéité masculine de la fin de l’Enéolithique tchèque. Ainsi, l’existence de sociétés campaniformes - et cordées - pratiquant l’exogamie dans l’aire orientale suffit-elle à expliquer la diffusion du Campaniforme? 218 De la discussion à l’interprétation 8.3. Une proposition de dynamique des peuplements campaniformes Les cinq occupations analysées offrent un portrait riche et complexe de la société du 3e millénaire avant notre ère. Il n’est décemment pas envisageable de s’attacher à une - et une seule - explication possible du Campaniforme à l’échelle de l’Europe. En effet, selon les territoires, différents comportements doivent être mis en évidence. Le rôle et l’implication du fonds local dans l’émergence du Campaniforme ne sont pas les mêmes selon les régions (figure 100). Seuls le nord de l’Espagne et la Bohême révèlent des liens étroits entre l’occupation campaniforme et leur substrat. Ce qui n’est pas incompatible avec les résultats des composantes archéologiques qui ont à plusieurs reprises défini ces deux régions comme des territoires importants et probablement à l’origine du phénomène, ou du moins d’un certain - voire de plusieurs Campaniforme. Figure 100 : Dynamique des peuplements campaniformes. Synthèse de l’implication du fonds régional dans la mise en place du Campaniforme. Pour les trois autres régions (sud de la France, Suisse et Hongrie), des influences populationnelles externes sont à l’origine de leur occupation campaniforme, mais leur apport n’est pas le même. Un renouvellement total - ou du moins un fort impact exogène - est manifeste pour le sud de la France et la Hongrie. En effet, les populations campaniformes se distinguent du substrat pour ces deux territoires de manière évidente. La Suisse, quant à elle, possède un fonds régional perturbé par un renouvellement partiel ou une intégration d’individus étrangers. Cette situation en territoire helvétique semble déjà présente, même si moins intensément, dès le Néolithique récent. La morphologie dentaire a permis de mettre en évidence deux sphères populationnelles sur le territoire étudié (figure 101). Les deux entités se distinguent clairement l’une de l’autre. Nous avons ainsi des Campaniformes méridionaux et des Campaniformes orientaux, que nous désignons respectivement par Bell Beakers (en référence à la céramique décorée qui semble originaire du bassin méditerranéen) et Beakers (en référence également au gobelet cordé). Nous sommes bel et bien en présence d’une sphère méridionale et d’une sphère orientale pour lesquelles le fonds populationnel n’est pas le même. Ce ne sont pas des traits particuliers qui les différencient, mais l’ensemble de la variabilité exprimée par la morphologie dentaire. 219 Chapitre 8 Figure 101 : Dynamique des peuplements campaniformes. Deux sphères populationnelles se distinguent. Les Campaniformes méridionaux, nord de l’Espagne et sud de la France, sont très proches et les populations helvétiques se rattachent fortement à leur morphologie. Ils forment un ensemble très homogène. Les Campaniformes orientaux - représentés ici par les populations de la Bohême - dévoilent une certaine cohésion qui paraît toutefois moins hermétique. Indépendamment de la problématique difficile de compréhension de la mise en place du Campaniforme sur un territoire aussi vaste, les données anthropologiques dentaires montrent qu’au sein des sphères populationnelles, les individus présentent des comportements clairement différents. Nous avons évoqué la possibilité que la diffusion du Campaniforme soit la conséquence d’une société pratiquant l’exogamie dans l’aire orientale. Pour ce faire, nous devons considérer que les producteurs des céramiques sont les femmes. Pouvons-nous exclure que les hommes soient les producteurs des céramiques ? Non, néanmoins si la production de la céramique est domestique [...] les études ethnoarchéologiques ont montré que, dans ce cas, la règle universelle est que les artisans sont des femmes. Les hommes ne sont potiers que dans le cadre d’une activité spécialisée et ils pratiquent alors leur métier dans des ateliers extérieurs à l’habitat familial ou en tant que potiers itinérants (Burri 2006, p. 119). Lors d’une union exogame, comment la femme productrice de la céramique s’adapte-t-elle face aux traditions céramiques d’origine et aux traditions nouvelles ? Des travaux sur les processus d’emprunts entre diverses traditions céramiques au Sénégal ont permis de mettre en évidence des éléments très intéressants (Gelbert 2003). En effet, si les techniques de façonnage nouvelles s’empruntent facilement, l’adoption d’une tradition céramique exogène ne signifie pas nécessairement une adaptation culturelle totale. De ce fait, une potière peut maintenir ou changer sa tradition si une valorisation est possible (E. Burri, communication personnelle). Ainsi, si la production de la céramique est domestique - ce qui semble le cas au Campaniforme (Salanova 1997, Besse à paraître) - par conséquent réalisée par les femmes, et si les potières ont adopté par emprunt les traditions méridionales, il est alors possible que le Campaniforme se soit diffusé au sein d’une sphère exogame dans l’aire orientale. 220 De la discussion à l’interprétation Sur la base de ces résultats est-il alors possible d’aborder un modèle de diffusion du Campaniforme ? Les figures 102 et 103 synthétisent la manière selon laquelle l’expansion pourrait avoir eu lieu. Au sein de la sphère méridionale : les Bell Beakers L’émergence du Campaniforme dans la sphère méridionale s’est produite par déplacements d’individus depuis la péninsule Ibérique. Les flèches pleines de la figure 102 indiquent le sens de l’expansion du phénomène mis en évidence par les traits non métriques. L’impact biologique s’enregistre au moins jusqu’en Suisse, voire même en Hongrie. Ainsi, les Bell Beakers - petits groupes d’individus munis de leur matériel et de leur savoir-faire - sont à l’origine de la diffusion du Campaniforme dans cette portion du phénomène. Les flèches en pointillé de la figure 102 reproduisent la propagation théorique des éléments méridionaux vers l’est. Figure 102 : Dynamique des peuplements campaniformes de la phase 1 (environ 3000 2500 av. J.-C.) : diffusion au sein de la sphère méridionale. Figure 103 : Dynamique des peuplements campaniformes de la phase 2 (environ 2500 2000 av. J.-C.) : diffusion au sein de la sphère orientale. 221 Chapitre 8 Au sein de la sphère orientale : les Beakers La situation dans la sphère orientale est plus complexe. Les données dentaires vont dans le sens d’une évolution au sein de la même société. Néanmoins, les femmes - cordées et campaniformes - se distinguent du fonds local, résultant probablement de sociétés pratiquant l’exogamie. Les flèches pleines de la figure 103 reproduisent schématiquement une possible diffusion du Campaniforme chez les Beakers. Les flèches en pointillé de la figure 103 concernent la propagation théorique des éléments orientaux vers l’ouest. Pour N. Brodie (2001), ce sont les femmes campaniformes orientales qui intègrent l’aire occidentale. Nos données n’appuient pas cette hypothèse, ni même l’inverse. En effet, les morphologies dentaires divergent fortement dans les deux sphères. Selon nos résultats, l’origine des femmes campaniformes du domaine oriental ne peut pas être attribuée aux Bell Beakers. Nous imaginons donc des mouvements opposés de sociétés exogames dans l’aire orientale. Ainsi, pour le Cordé, la diffusion de la culture se développe en direction de l’ouest. En revanche, l’expansion du Campaniforme - issu lui-même de la société cordée - se déroule vers l’est du domaine. Nous ne pouvons toutefois pas exclure - actuellement - l’existence d’un apport populationnel des Bell Beakers, ni le mesurer. En conséquence et sur la base des datations radiocarbone actuellement disponibles, mettant en évidence un gradient sud-ouest - nord-est dans la mise en place du Campaniforme, nous proposons un modèle de diffusion en deux temps : Phase 1. Des groupes d’individus Bell Beakers migrent depuis la péninsule Ibérique vers l’est, parallèlement l’aire orientale est occupée par la culture Cordé (et ce jusqu’en Suisse orientale). Phase 2. Une partie de la société Cordé à la limite du phénomène s’individualise et adopte, par emprunt, une partie des traditions méridionales campaniformes. Cette nouvelle société - les Beakers - se propage vers l’est. En même temps, certains éléments orientaux se diffusent à l’ouest. 222 Conclusions Et bien oui, encore un regard nouveau posé sur le Campaniforme ! Entité de la fin du Néolithique européen, le Campaniforme, que l’on a du mal à définir et à cerner comme un tout, a bénéficié d’interprétations diverses et variées. A l’origine de ces difficultés, son ample répartition géographique et ses composantes culturelles complexes. En effet, il s’étend des îles britanniques aux côtes de l’Afrique du Nord et des côtes atlantiques à la Hongrie, et le noeud du problème [...], mais qui en fait aussi tout l’intérêt, est la présence conjointe d’éléments paneuropéens et d’éléments régionaux (Besse et al. 2007, p. 193). Depuis quelques années maintenant, la recherche au sein du laboratoire d’archéologie préhistorique et d’histoire des peuplements de l’Université de Genève privilégie les études à l’échelle du phénomène. Et notre travail, intégré au sein d’un programme interdisciplinaire cherchant à identifier l’implication du fonds local dans la mise en place du Campaniforme, concerne l’étude des protagonistes à qui l’on doit cette énigme. Ainsi notre étude a poursuivi l’objectif de déterminer si l’émergence du Campaniforme coïncidait ou non avec un renouvellement de la population. Afin d’y répondre : • Cinq expressions différentes de l’occupation campaniforme européenne ont été sélectionnées. L’aire orientale est représentée par la Bohême et par la Hongrie. L’aire méridionale est constituée par le sud de la France et le nord de l’Espagne. Enfin la dernière région étudiée - la Suisse - se situe à l’articulation entre les deux domaines. • L’observation des traits non métriques dentaires a été retenue. Nous avons montré que ces variables sont de bons indicateurs des distances entre populations en nous attachant à leur valeur biologique, à la fiabilité du système d’enregistrement et au potentiel observé lors d’études actualistes. Nous avons eu à notre disposition près de 2’000 individus appartenant, non seulement au Campaniforme, mais également - et selon les régions - aux diverses cultures pré et post campaniformes. Nous avons examiné 111 variables, enregistrées sur les dentitions permanente et déciduale, représentant 530 observations possibles par individu. Nous avons donc enregistré 255'000 données. La fiabilité du système d’enregistrement (détermination des dents et des traits non métriques) a été testée et notre faible discordance a permis la validation de nos données. Un traitement préliminaire (vérification de la transcription, élimination des données non pertinentes, traitement des données bilatérales, évaluation des relations et du dimorphisme sexuel des variables, appréciation des effectifs des échantillons) a servi à l’épuration des données afin de ne garder que les informations mobilisables au sein des analyses. Nous avons finalement réalisé une série d’analyses multivariées (des analyses ascendantes hiérarchiques complétées par des bootstrap, des multidimensional scaling complétés par des minimal spanning tree et des analyses en composantes principales) à plusieurs niveaux de différenciation (échelles intra- et interrégionales, temporelle et sexes séparés). A l’issue des analyses, nous avons pu montrer que : • Le fonds local n’est pas impliqué dans la mise en place du Campaniforme sur l’ensemble des régions étudiées. En effet, une continuité de peuplement est clairement constatée pour le nord de l’Espagne et la Bohême. Un renouvellement de la population est en revanche identifié pour le sud de la France, la Hongrie et la Suisse - quoique pour cette dernière, il ne semble que partiel. • Deux sphères populationnelles campaniformes distinctes ont été reconnues. Elles correspondent aux aires orientale et méridionale du phénomène. Des populations différentes sont donc présentes : les Campaniformes méridionaux - les Bell Beakers - et les Campaniformes orientaux - les Beakers. 223 • Les comportements au sein des sphères populationnelles sont totalement différents. Les Bell Beakers forment un ensemble très homogène et fermé, et les Beakers constituent un groupe cohérent quoique moins hermétique. A l’aide de ces éléments nous avons proposé un scénario des peuplements campaniformes à l’échelle des domaines étudiés : Phase 1 Dans la sphère méridionale, des petits groupes de Bell Beakers migrent depuis la péninsule Ibérique vers l’est. Leur impact populationnel s’identifie au moins jusqu’en Suisse. L’aire orientale est peuplée par les cordés. Phase 2 Dans la sphère orientale, une partie de la communauté cordée se particularise en adoptant les traditions méridionales apportées par les Bell Beakers jusqu’aux portes de l’aire cordée. Cette nouvelle société - les Beakers - colonise l’est dans un système d’exogamie diffusant ces nouvelles traditions. Des éléments orientaux se diffusent également vers l’ouest. Enfin, ce contact entre les deux communautés - les Bell Beakers et les Beakers - donne naissance à l’apparente uniformité du Campaniforme. Notre étude n’a pas la prétention d’avoir résolu l’énigme posée par le Campaniforme à l’échelle du phénomène. Néanmoins elle offre des éléments de compréhension intéressants. Il n’est plus envisageable aujourd’hui de considérer le Campaniforme comme la résultante d’un événement unique. Deux fonds populationnels différents ont été mis en évidence au sein desquels les modalités de diffusion du Campaniforme ne sont pas les mêmes. La forte cohésion des populations campaniformes dans l’aire méridionale plaide en faveur d’un apport de sang nouveau par la migration de groupes d’individus. En revanche, les femmes sont le vecteur de diffusion du complexe dans la sphère orientale. L’impact de ces différents comportements n’est pas le même partout, certaines régions voient le cours de leur histoire profondément modifié par l’irruption du Campaniforme, d’autres semblent le rejeter (Salanova et Sohn 2007, p. 84). Les perspectives véhiculées par notre étude sont multiples et se prolongent à travers divers axes de recherches. Dans un premier temps, il faudrait compléter l’étude : • Dans la sphère méridionale (notamment Portugal et Catalogne). Etendre l’analyse à l’ensemble de la péninsule Ibérique devrait valider l’implication du fonds local dans la mise en place du Campaniforme. • Dans la sphère orientale (Allemagne, Autriche, Pologne et Moravie). Poursuivre l’étude de ce domaine consentirait, non seulement à identifier le prolongement de l’impact des Bell Beakers dans l’aire orientale, mais également à préciser la situation originale de la Hongrie, où l’on soupçonne un certain apport populationnel méridional. Dans un deuxième temps, il s’agirait de développer l’étude à l’échelle du phénomène : • L’aire septentrionale. Aborder ce domaine n’est pas chose aisée. Néanmoins, cela permettrait, à n’en pas douter, une meilleure compréhension de la dynamique des peuplements campaniformes sur l’ensemble du phénomène. • L’occupation campaniforme italienne. Le Campaniforme italien est complexe et s’en approcher par l’anthropologie biologique s’avère difficile sur la péninsule puisque les vestiges osseux ne sont généralement pas conservés. En revanche, la Sardaigne et la Sicile sont très bien documentées et fourniraient des indications des plus intéressantes sur la diffusion et l’implantation insulaires du Campaniforme en zone méridionale. 224 Conclusions Enfin, il faudrait vérifier et quantifier, dans les différentes régions analysées, l’apport de populations externes à l’aide des analyses chimiques des ossements ou encore de l’ADN ancien, ce que la morphologie dentaire seule n’est pas actuellement en mesure de réaliser. Nous l’avons donc bien constaté dans cette étude, les individus associés au Campaniforme ont encore bien des choses à raconter sur leur histoire. Des éléments nouveaux, ou de simples précisions, ouvrent la voie à une meilleure compréhension des comportements à l’origine de l’émergence de cette énigme, de ce phénomène ou encore de cette question de la fin du Néolithique européen ! 225 De laboratoires en instituts : quelques souvenirs en images... Palencia, Espagne Prague, Tchéquie Valladolid, Espagne Bordeaux, France Szeged, Hongrie Szeged, Hongrie Szeged, Hongrie Prague, Tchéquie Marseille, France Pot-pourri d’ici et là... Marseille, France 227 Bibliographie Aas (I.H.), Risnes (S.). 1979a. The depth of the lingual fossa in the permanent incisors of Norwegians. I. Method of measurement, statistical distribution and sex dimorphism. American Journal of Physical Anthropology 50, 335-340. Aas (I.H.), Risnes (S.). 1979b. The depth of the lingual fossa in the permanent incisors of Norwegians. II. Method of measurement, statistical distribution and sex dimorphism. American Journal of Physical Anthropology 50, 341-348. About (I.), Bottero (M.-J.), Denato (P. de), Camps (J.), Franquin (J.-C.), Mitsiadis (T.A.). 2000. Human Dentin Production in Vitro. Experimental Cell Research 258, 33-41. About (I.), Camps (J.), Mitsiadis (T.A.), Bottero (M.-J.), Butler (W.), Franquin (J.-C.). 2002. Influence of resinous monomers on the differentiation in vitro of human pulp cells into odontoblasts. Journal of Biomedical Materials Research (Applied Biomaterials) 63, 418-423. Alcade Crespo, (G.Y.), Rincon Vila (R.). 1986. El conjunto funerario de Cueva Tino. La Horadada, Mave (Palencia). Palencia: Tello Téllez de Meneses. Alexandersen (V.). 1962. Root conditions in human lower canines with special regard to double-rooted canines. II. Occurrence of double-rooted lower canines in Homo sapiens and other primates. Saetryk af Tandlaegebladet 66, 729-760. Alt (K.W.) 1997. Odontologische Verwandtschaft analyse: Individuelle Charakteristika der Zähne in ihrer Bedeutung für Anthropologie, Archäologie und Rechtsmedizin. Stuttgart: G. Fischer. Alt (K.W.). 2005. Informe antropologico de los enterramientos campaniformes del Tumulo de la Sima. In: Rojo Guerra (M.A.), Kunst (M.), Garrido Pena (R.), Garcia Martinez De Lagran (I.), ed. Un Desafío a la Eternidad. Tumbas Monumentales del Valle de Ambrona (Soria, España). Junta de Castilla y León: Monografías. Arqueologia en Castilla y Leon, Memorias 14, 322. Alt (K.W.), Vach (W.). 1991. The reconstruction of “genetic kindship” in prehistoric burial complexes: Problems and statistics. In: Bock (H.H.), Ihm (P.), ed. Classification, data analysis and knowledge organisation. Heidelberg: Springer, 299-310. Alt (K.W.), Vach (W.). 1992. Non-spatial analysis of “genetic kindship” in skeletal remains. In: Schader (M.), ed. Analyzing and modelling data and knowledge organisation. Berlin: Springer, 247-256. Alt (K.W.), Vach (W.). 1995. Odontologic kinship analysis in skeletal remains: Concepts, methods and results. Forensic Science International 74, 99-113. Alt (K.W.), Türp (J.C.). 1997. Dental anthropology. In: Alt (K.W.), Brace (C.L.), Türp (J.C.), ed. The history of dental anthropology. Stuttgart: G. Fischer, 15-39. Alt (K.W.), Pichler (S.), Vach (W.). 1998. Dental traits for population studies: the Freiburg University Dental Trait System (FU-DTS). Freiburg: Freiburg University (Catalogue non publié). Alvesalo (L.), Nuutila (M.), Portin (P.). 1975. The cusp of Carabelli occurrence in the first upper molars and evaluation of its heritability. Acta Odontologica Scandinavica 33, 191-197. Amrane (Y.), Ducreux (F.). 1996. La déviation de la RN 73 à Saint-Marcel. Sauvetage des sites de la Noue et sur le Bief. Service régional de l'Archéologie de Bourgogne (Document synthèse non publié). Andres (M.T.), Garcia (M.L.), Sesma (J.). 1983. Una tumba destruida por el fuego: el sepulcro campaniforme de Tres Montes en Las Bardenas Reales (Navarra). In: Rojo Guerra (M.A.), Kunst (M.), ed. Sobre el significado del fuego en los rituales funerarios del Neolitico. Valladolid: Universidad. Studia archaeologica 91, 191-218. Ash (M.), ed. 1993. Wheeler’s dental anatomy, physiology, and occlusion. London: W.B. Saunders, 7e ed, 1940. Aspes (A.), Fasani (L.), Gaggia (F.), ed. 1980. La preistoria del Lago di Garda. Catalogo della mostra (Verona, 8 aprile-30 settembre 1980). Verona: Museo Civico di Storia Naturale. Aubry (M.) 2003. Etude paléo-odontologique de populations préhistoriques et historiques de Provence: L’hypogée chalcolithique de Roaix (Vaucluse) et le cimetière de Saint-Pierre de l’Almanarre. Université de la Méditerranée (Aix-Marseille II): Laboratoire d’anthropologie, secteur Nord (Thèse de doctorat non publiée). Aubry (M.), Mafart (B.), Donat (B.), Brau (J.J.). 2003. Brief communication: Study of noncarious cervical tooth lesions in samples of prehistoric, historic, and modern populations from the South of France. American Journal of Physical Anthropology 121, 10-14. Axelsson (G.), Kirveskari (P.). 1982. Correlation between lower molar occlusal traits in Icelanders. In: Kurten (B.), ed. Teeth: form, function, evolution. New York: Columbia University Press, 237-244. 229 Axelsson (S.), Bjornland (T.), Kjaer (I.), Heiberg (A.), Storhaug (K.). 2003. Dental characteristics in Williams syndrome: a clinical and radiographic evaluation. Acta Odontologica Scandinavica 61, 129-36. Axenovich (T.I.), Zorkoltsova (I.V.), Aulchenko (Yu.S.), Knyazev (S.P.), Kulikova (A.V.). 2004. Inheritance of hypodontia in Kerry Blue Terrier dogs. Russian Journal of Genetics 40, 529-536. Bader (R.S.). 1965. Heritability of Dental Characters in the House Mouse. Evolution 19, 378-384. Bailit (H.L.), Sung (B.). 1968. Maternal effects on the developing dentition. Archives of Oral Biology 13, 155-161. Bailit (H.L.), Workman (P.L.), Niswander (J.D.), MacLean (C.J.). 1970. Dental asymmetry as an indicator of genetic and environmental conditions in human populations. Human Biology 42, 626-638. Bailly (M.). 2002. La flèche et l’éclat. Production et consommation des outillages lithiques taillés du Néolithique final à l’âge du Bronze ancien entre Saône et Rhône (2600-2200 av. J.-C.). Besançon: Université de FrancheComté (Thèse de doctorat non publiée). Bailly (M.), Salanova (L.). 1999. Les dates radiocarbone du Campaniforme en Europe occidentale: analyse critique des principales séries de dates. In: Evin (J.), Oberlin (C.), et al., ed. C14 et Archéologie. 3e Congrès international (Lyon, 6-10 avril 1998). Mémoires de la Société Préhistorique Française 26, 219-224. Barfield (L.H.). 1983. The Chalcolitic cemetery at Manerba del Garda. Antiquity 57, 220, 116-123. Barge-Mahieu (H.). 1992. L'habitat des Calades à Orgon (Bouches-du-Rhône). In: Barge-Mahieu (H.), ed. Le Campaniforme dans le Midi de la France: origine et identité culturelle. Exposition (Orgon, 6 juillet-28 septembre 1992). Marseille: Etudes et Prospectives Archéologiques, 22-30. Bartelheim (M.). 1998. Studien zur böhmischen Aunjetitzer Kultur: chronologische und chorologische Untersuchungen. Bonn: R. Habelt. Universitätsforschungen zur prähistorischen Archäologie 46. Bath-Bílková (B.).1973. K problému původu hřiven-Zur Herkunftsfrage des Halsingbarren. Památky Archeologické 64, 24-41. Baudais (D.), Kramar (C.). 1990. La nécropole néolithique de Corseaux "en Seyton" (VD, Suisse): archéologie et anthropologie. Lausanne: Bibliothèque historique vaudoise. Cahiers d’Archéologie Romande 51-Document du Département d’anthropologie et d’écologie de l’Université de Genève 15. Baudais (D.), Brunier (C.), Curdy (P.), David-Elbiali (M.), Favre (S.), Gallay (A.), Moinat (P.), Mottet (M.), Voruz (J.-L.), Winiger (A.). 1989-1990. Le Néolithique de la région de Sion (Valais): un bilan. Bulletin du Centre Genevois d’Anthropologie 2, 5-56. Baume (R.M.), Crawford (M.H.). 1980. Discrete dental trait asymmetry in Mexican and Belizean groups. American Journal of Physical Anthropology 52, 315-321. Bello (S.), Thomann (A.), Rabino Massa (E.), Dutour (O.). 2003. Quantification de l’état de conservation des collections ostéoarchéologiques et ses champs d’application en anthropologie. Antropo 5, 21-37. Beneš (J.), Dobeš (M.). 1992. Eine schnurkeramische Gräbergruppe und ein Objekt der Kugelamphorenkultur aus Hrdlovka (NW Böhmen). In: Buchvaldek (M.), Strahm (C.), ed. Die kontinentaleuropäischen Gruppen der Kultur mit Schnurkeramik (Schnurkeramik symposium 1990). Praehistorica 19, 67-79. Benz (M.), Strahm (C.), Van Willigen (S.). 1998. Le campaniforme: phénomène et culture archéologique. Bulletin de la Société Préhistorique Française 95, 305-314. Bermond Montanari (G.). 1998. Gli insedamenti campaniformi di Rubiera e Sant'Ilario d'Enza. In: Nicolis (F.), Mottes (E.), ed. Simbolo ed enigma: il bicchiere campaniforme e l'Italia nella preistoria europea del III millennio a.C. Catalogo della mostra (Riva del Garda, 1998). Trento: Servizio Beni Culturali, Provincia Autonoma, 92-94. Bermond Montanari (G.), Cremaschi (M.), Sala (B.). 1982. Rubiera: insediamento del Vaso campaniforme. Preistoria alpina 18, 79-109. Berry (A.C.). 1976. Anthropological value of minor variants of the dental crown. American Journal of Physical Anthropology 45, 257-268. Berry (A.C.). 1978. Anthropological and family studies on minor variants of the dental crown. In: Butler (P.M.), Joysey (K.A.), ed. Development, function and evolution of teeth. New-York: Academic Press, 81-98. Berry (R.J.). 1963. Epigenetic polymorphism in wild populations of Mus Musculus. Genetical Research 4, 193-220. Berry (R.J.), Berry (A.C.). 1967. Epigenetic variation in the human cranium. Journal of Anatomy 101, 361-379. Bertemes (F.), Heyd (V.). 1996. Définition et origine de l'âge du Bronze ancien en Europe centrale. In: Mordant (C.), Gaiffe (O.), ed. Cultures et sociétés du Bronze ancien en Europe. Colloque sur les Fondements culturels, techniques, économiques et sociaux des débuts de l'âge du Bronze. Congrès national des Sociétés savantes, 117, section de pré- et protohistoire (Clermond-Ferrand, 27-29 octobre 1992). Paris: Editions du Comité des travaux historiques et scientifiques (C.T.H.S.), 13-36. Besse (M.). 1998a. L'apport de l'étude de la céramique commune à la compréhension du Campaniforme. Bulletin de la Société Préhistorique Française 95, 327-331. 230 Bibliographie Besse (M.). 1998b. The Bell-Beaker culture in the Switzerland: documentary assessment and an attempt to synthesize. In: Section 10: Âge du Cuivre au Proche-Orient et en Europe. International congress of prehistoric and protohistoric sciences, 13 (Forlì, 8-14 septembre 1996): volume 4. Forlì: A.B.A.C.O, 117126. Besse (M.). 2001. L'Europe continentale, la région Rhin-Rhône et l'habitat de Derrière-le-Château (Ain-France): la céramique commune campaniforme. Université de Genève: Département d’anthropologie et d’écologie (Thèse de doctorat non publiée). Besse (M.). 2003. Les céramiques communes des Campaniformes européens. Gallia Préhistoire 45, 205-258. Besse (M.). 2004a. Bell Beaker common ware during the third millennium BC in Europe. In: Czebreszuk (J.), ed. Similar but different: Bell Beakers in Europe. Poznań: Adam Mickiewicz University, Institute of Prehistory, 127-148. Besse (M.). 2004b. Des Campaniformes européens au Campaniforme méditerranéen. Bulletin de la Société Préhistorique Française 101, 215-222. Besse (M.). 2006. Les archers néolithiques en Pays de Neuchâtel. In: Arnold (B.), Bauermeister (N.), Ramseyer (D.), ed. Archéologie plurielle: mélanges offerts à Michel Egloff à l'occasion de son 65e anniversaire. Hauterive: Service et Musée cantonal d'archéologie de Neuchâtel. Archéologie Neuchâteloise 34, 37-45. Besse (M.). (à paraître). Le Campaniforme de Rances. In: El Biali David (M.), ed. Archéologie et préhistoire à Rances (Vaud, Suisse). Besse (M.), Desideri (J.). 2005. Bell Beaker diversity: settlements, burials and ceramics-La diversidad Campaniforme: Hábitats, sepulturas y cerámicas. In: Rojo Guerra (M.A.), Garrido Pena (R.), Garcia Martinez De Lagran (I.), ed. El Campaniforme en la Peninsula Ibérica y su contexto europeo-Bell Beakers in the Iberian Peninsula and their european context. Valladolid: Universidad de Valladolid. Arte y Arqueologia 21, 61-106. Besse (M.), Desideri (J.), Piguet (M.). (2007). Céramiques, sociétés et histoire des peuplements dans l’Europe campaniforme. In: Guilaine (J.), ed. Le Chalcolithique et la construction des inégalités. Tome I: le continent européen. Séminaires du Collège de France. Paris: Editions Errance (Collection des Hespérides), 191-209. Biggerstaff (R.H.). 1973. Heritability of Carabelli’s cusp in twins. Journal of Dental Research 52, 40-44. Biggerstaff (R.H.). 1979. The biology of dental genetics. Yearbook of Physical Anthropology 22, 215-227. Blajerová (M.). 1957. Anthropologický posudek-Tuchoměřice (viz Šneidrová K.). Archeologické Rozhledy 9, 414415. Blajerová (M.). 1958. Anthropologický posudek-Sulejovice (viz Moucha V.). Archeologické Rozhledy 10, 77-78. Blajerová (M.). 1962. Antropologické Zhodnoceni Mladoeneolitickych Kostrovych Pohřbů Z Libochovic, Zabovřesk A Stětí. Sborník Severočeského musea. Historia 3, 163-166. Blanco (R.), Chakraborty (R.). 1977. The genetics of shovel shape in maxillary central incisors in man. American Journal of Physical Anthropology 44, 233-236. Blasco (C.), Baena (J.), Recuero (V.). 1994. Los asentamientos. In: Blasco (C.), ed. El horizonte campaniforme de la region de Madrid en el centenario de Ciempozuelos. Madrid: Universidad autonoma, 47-74. Boas (F.). 1912. Changes in bodily form of descendants of immigrants. New York: Columbia University Press. Bocksberger (O.-J.). 1976. Le dolmen M VI: texte, catalogue et planches. Lausanne: Bibliothèque historique vaudoise. Cahiers d’Archéologie Romande 6 et 7. Bocksberger (O.-J.). 1978. Horizon supérieur: secteur occidental et tombes Bronze ancien: texte, catalogue et planches. Lausanne: Bibliothèque historique vaudoise. Cahiers d’Archéologie Romande 13 et 14. Boklage (C.E.). 1987. Developmental differences between singletons ans twins in distributions of dental diameter asymmetries. American Journal of Physical Anthropology 74, 319-331. Boraas (J.C.), Messer (L.B.), Till (M.J.). 1988. A genetic contribution to dental caries, occlusion, and morphology as demonstrated by twins reared apart. Journal of Dental Research 67, 1150-1155. Bosch-Gimpera (P.). 1926. Glockenbecherkultur. In: Ebert (M.), ed. Reallexikon der Vorgeschichte (Berlin) 4, 344362. Bouchet (J.-M.), Burnez (C.), Fouéré (P.). 1990. La Grande Pigouille à Belluire (Charente-Maritime). Bulletin de la Société Préhistorique Française 87, 153-160. Bouville (C.) 1971. Les reste humains du dolmen des Peyraoutes (Roquefort-les-Pins, Alpes-Maritimes). Contribution à l’étude des populations du Néolithique et de l’âge du Bronze en Provence. Université de Provence: Géologie historique et paléontologie (Thèse de doctorat non publiée). Bouville (C.) 1980. L’hypogée chalcolithique de Roaix. Apport à l’étude de la démographie en Provence. Bulletins et Mémoires de la Société d’Anthropologie de Paris 7, 85–89. 231 Brabant (H.). 1967. Comparison of the characteristics and anomalies of the deciduous and permanent dentition. Journal of Dental Research 46, 897-902. Brewer-Carias (C.A.), Le Blanc (S.), Neel (J.V.). 1976. Genetic structure of a tribal population: the Yanomama Indians. XIII: Dental microdifferentiation. American Journal of Physical Anthropology 44, 5-14. Brodie (N.). 1994. The Neolithic-Bronze Age: transition in Britain: a critical review of some archaeological and craniological concepts. Oxford: Tempus Reparatum. BAR: British series 238. Brodie (N.). 2001. Technological frontiers and the emergence of the Beaker culture. In: Nicolis (F.), ed. Bell Beakers today: pottery, people, culture, symbols in prehistoric Europe: volume 2. International Colloquium (Riva del Garda, Trento, 11-16 May 1998). Trento: Servizio Beni Culturali, Provincia Autonoma di Trento, 111-124. Brothwell (D.R.), Carbonell (V.M.), Goose (D.H.). 1963. Congenital absence of teeth in human populations. In: Brothwell (D.R.), ed. Dental anthropology. London: Pergammon Press, 179-190. Brook (A.H.). 1984. A unifying aetiological explanation for anomalies of human tooth number and size. Archives of Oral Biology 29, 373-378. Buchvaldek (M.). 1967. Die Schnurkeramik in Böhmen. Praha: University Karlova (Acta Universitatis Carolinae). Buchvaldek (M.). 1978. Otázka kontinuity v českomarvském mladśim eneolitu-Zur Frage der Kontinuität im jüngeren Äneolithikum in Böhmen und Mähren. Praha: University Karlova. Praehistorica 7, Varia Archeologica 1, 35-64. Buchvaldek (M.). 1985. Kultura se śnůrovou keramikou ve strědni Evropě: I. Skupiny mezi Harcem a Bilýmí Karpaty. Praha: University Karlova. Praehistorica 12, 1-160. Buchvaldek (M.). 1986. Zum gemeineuropaïschen Horizont der Schnurkeramik. Praehistorische Zeitung 61, 129150. Buchvaldek (M.). 1990. Pohřebiště lidu se zvoncovitými poháry. Ein Gräberfeld der Glockenbecherkultur. In: Lochenice, Z archeologických výzkumů na katastru obce. Praha: University Karlova. Praehistorica 16, 29-50. Buchvaldek (M.), Koutecký (D.). 1972. Interpretation des schnurkeramischen Gräberfeldes von Vikletice. Památky Archeologické 63, 142-179. Buchvaldek (M.), Popelka (M.). 1994. Poznámky k pohřebišti se šňůrovou keramikou v Praze-JinonicíchBemerkungen zum schnurkeramischen Gräberfeld in Prag-Jinonice. Praha: University Karlova. Praehistorica 21, Varia Archeologica 6, 21-52. Buchvaldek (M.), Vencl (S.). 1975. Eneolitický etážovitý džbánek z Mochova, okr. Praha-východ-An Aeneolithic Storied Jar from Mochov, District Praha-East. Archeologické Rozhledy 27, 121-125. Buchvaldek (M.), Moucha (V.), Popelka (M.), Vojtěchovská (I.). 1997. Katalog šňůrové keramiky v Čechách XIXIV. Kladensko, Slánsko, Kralupsko a Praha-západ-Die Kataloge der Schnurkeramik in Böhmen XI.XIV. Die Räume von Kladno, Slaný, Kralupy na Vltavou, Praha-West. Praha: University Karlova. Praehistorica 22, Varia Archeologica 7, 113-256. Bueno Ramírez (P.), Balbín Behrmann (R.), Barroso Bermejo (R.). 2000. Valle de las Higueras (Huecas, Toledo, Espana). Una necrópolis Ciempozuelos con cuevas artificiales al interior de la Península. VISEU: Centro de Estudos Pre-Historicos da Beira Alta 8, 49-80. Bueno Ramírez (P.), Barroso Bermejo (R.), Balbín Behrmann (R.). 2005. Ritual campaniforme, ritual colectivo: la necrópolis de cuevas artificiales del Valle de las Higueras, Huecas, Toledo. Trabajos de Prehistoria 62, 6790. Burnez (C.), Lagarde (M.-C.). 1986. Campaniforme sur le site des Loups à Echiré (Deux-Sèvres). Groupe Vendéen d'Etudes Préhistoriques, numéro spécial, 97-100. Burri (E.). 2006. Les stations du Néolithique moyen de Concise (Vaud, Suisse): céramique, analyses spatiales et interprétations en termes d'histoire des peuplements. Genève: Université de Genève: Département d’anthropologie et d’écologie (Thèse de doctorat non publiée). Butler (P.M.) 1939. Studies of the mammalian dentition: Differentiation of the postcanine dentition. Proceedings of the Zoological Society of London 109, 1-36. Butler (P.M.). 1978. Molar cusp nomenclature and homology. In: Butler (P.M.), Joysey (K.A.), ed. Development, Function and Evolution of Teeth. New York: Academic Press, 439-452. Butler (P.M.). 2001. What happened to the Field theory. In: Brook (A.), ed. Dental morphology 2001. Proceedings of the 12th International Symposium on Dental Morphology. Sheffield: Academic Press Ltd, 3-12. Campbell (N.A.), Reece (J.B.). 2004. Biologie. Bruxelles: De Boeck Université. Carbonell (V.M.), Goose (D.H.). 1963. Congenital absence of teeth in human populations. In: Brothwell (D.R.), ed. Dental anthropology. London: Pergammon Press, 179-190. Cardoso (J.L.). 1989. Leceia. Resultados des Escavaçoes Realizadas em 1983-1988. Camara municipal de Oeiras. 232 Bibliographie Cardoso (J.L.). 2001. Le phénomène campaniforme dans les basses vallées du Tage et Sado (Portugal). In: Nicolis (F.), ed. Bell Beakers today: pottery, people, culture, symbols in prehistoric Europe: volume 1. International Colloquium (Riva del Garda, Trento, 11-16 May 1998). Trento: Servizio Beni Culturali, Provincia Autonoma di Trento, 139-154. Cardoso (J.L.). 2004. A baixa estremadura dos finais do IV milénio A.C. até à chegada dos romanos: um ensaio de historia regional. Oeiras: Camara municipal. Estudios Arqueologicos de Oeiras 12. Carlsen (O.), Alexandersen (V.). 1999. Radix paramolaris and radix distomolaris in Danish permanent maxillary molars. Acta Odontologica Scandinavica 57, 283-289. Carlsen (O.), Alexandersen (V.). 2000. Radix mesiolingualis and radix distolingualis in a collection of permanent maxillary molars. Acta Odontologica Scandinavica 58, 229-236. Carson (E.A.). 2006. Heritability of cranial nonmetric traits. Human Biology 78, 383-402. Cartailhac (E.). 1886. Les âges préhistoriques de l'Espagne et du Portugal. Paris: Ch. Reinwald. Cattin (F.). (en cours). La métallurgie du cuivre dans les Alpes occidentales au 3e millénaire avant J.-C. Université de Genève: Département d’anthropologie et d’écologie (Thèse de doctorat). Cavalli-Sforza (L.L.), Menozzi (P.), Piazza (A.). 1994. The history and geography of human genes. Princeton, New Jersey: Princeton University Press. Ceccanti (M.), Cocchi (D.), Fornaciari (G.). 1981. Una grotticella sepolcrale eneolitica con vaso campaniforme scoperta presso Vecchiano (Pisa). Atti della Società Toscana di Scienze Naturali A 87, 375-391. Čech (P.), Černý (V.). 1996. Nové možnosti studia pohřebního ritu na příkladu únětických hrobů z Konobrží, okr. Most-Neue Möglichkeiten zum Studium des Bestattungsritus am Beispiel der Aunjetitzer Gräber aus Konobrže, Bez. Most (NW-Böhmen). Archeologické Rozhledy 48, 35-59. Černý (V.). 1996. Cranial variation of Eneolithic and Bronze Age people in Bohemia. Anthropologie 34, 333-341. Černý (V.). 2000. Velké Přílepy Antropologická analýza pohřebiště kultury se zvoncovitými poháry z Velkých Přílep-Kamýku-Physical anthropological analysis of a Bell Beaker culture cemetery from Velké PřílepyKamýk. Archeologie ve středních Čechách 4/1, Praha 79-84. Černý (V.), Veleminsky (P.). 1998. Die Anthropologie des Neolithikums in Böhmen. In: Bestattungsritus des böhmischen Neolithikums (5500-4200 B. C.). Praha: ARÚ AVČR, 145-158. Chagula (W.K.). 1960. The cusps on the mandibular molars of East Africans. American Journal of Physical Anthropology 18, 83-90. Chiaradia (M.), Gallay (A.), Todt (W.). 2003. Differencial contamination styles of prehistoric human teeth at a Swiss necropolis (Sion, Valais) inferred from lead and strontium isotopes. Applied geochemistry 18, 353370. Childe (V.G.). 1925. The dawn of European civilization, London: Routledge & P. Kegan. Chochol (J.). 1954. Anthropologické zhodnocení nálezu z Blšan-Analyse anthropologique de la trouvaille de Blšany. Archeologické Rozhledy 6, 745. Chochol (J.). 1955. Kostry z únětické jámy v Klučově-Squelettes de la fosse ouniétitzienne de Klučov, analyse anthropologique. Archeologické Rozhledy 7, 592-594. Chochol (J.). 1957. Kostra s trepanovanou lebkou z hrobu se šňůrovou keramikou v Kněževsi-Skelett mit trepaniertem Schädel aus enem schnurkeramischen Grabe in Kněževes. Archeologické Rozhledy 9, 713-714. Chochol (J.). 1958. Výsledky anthropologického zpracováni kostrových pozůstatků z Hrušova-Das Aunjetitzer Gräberfeld in Hrušov (Bez. Ml. Boleslav), Böhmen. Archeologické Rozhledy 10, 309-311. Chochol (J.). 1966. Antropologický příspěvek o obyvatelích Staré Kouřimě. In: Šolle (M.), ed. Stará Kouřim a projevy velkomoravské hmotné kultury v Čechách. Monumenta Archaeologica 15, 281-288. Chochol (J.). 1968. Kostry ze šňůrového pohřebiště v Polákách-Die Skelette aus dem schnurkeramischen Gräberfeld in Poláky (viz Koutecký D.). Archeologické Rozhledy 20, 291-293. Chochol (J.). 1976. Antropologické zhodnocení kosterných nálezů ze Širokých Třebčic-Anthropologische Untersuchung an Knochenfunden aus Široké Třebčice (viz Lehečková E.-Bubeník J.). Archeologické Rozhledy 28, 367-368. Chochol (J.). 1979a. Antropologické zhodnocení kosterných nálezů z hrobů šňůrové kultury v Konobržích, Obrnicích a Patokryjích-Anthropologische Auswertung von Skelettfunden aus Gräbern der Schnurkeramik in Konobrže, Obrnice und Patokryje. Archeologické Rozhledy 31, 23-26. Chochol (J.). 1979b. Antropologické určení kosterních pozůstatků z hrobů 75 a 77 v Polákách-The Anthropological Identification of Skeleton Remains from Graves 75, 77 (Migration Period) at Poláky (viz Smrž, Z.). Archeologické Rozhledy 31, 612-613. 233 Chochol (J.). 1980a. Antropologické zhodnocení kosterných nálezů únětické kultury z Prahy-ČimicAnthropologische Bewertung von Skeletüberresten der Úněticer Kultur aus Prag-Čimice. Archeologica Pragensia 1, 155-156. Chochol (J.). 1980b. Antropologické zhodnocení kosterných nálezů z hrobů KZP v Praze-AnthropologischeAuswertung von Skelettfunden der Glockenbecherkultur aus Prag-Kobylisy und Prag-Lipence. Archeologické Rozhledy 32, 132-133. Chochol (J.). 1980c. Antropologický rozbor pohřebiště šňůrové a únětické kultury v Praze-Čakovicích. Archaeologica Pragensia 1, 155-166. Chochol (J.). 1981a. Antropologické zhodnocení mladoúnětických koster z Chotěbudic-Anthropological evaluation of the Late Únětician skeletons from Chotěbudice (viz Rada I.). Archeologické Rozhledy 33, 634-635. Chochol (J.). 1981b. Antropologické zhodnocení kostry z hrobu LX v Plotištich n. L.-Die anthropologische Beurteilung des Skellets aus dem Grab LX in Plotiště n. L.. Archeologické Rozhledy 33, 485-486. Christian (J.C.), Kang (K.W.), Norton (J.A.). 1974. Choice of an estimate of genetic variance from twin data. American Journal of Human Genetics 26, 154-161. Claivaz-Caruzzo (D.). 1975. Anthropologie du dolmen néolithique MXI, site du Petit-Chasseur, Sion, Valais. Université de Genève: Département d’anthropologie et d’écologie (Travail de diplôme non publié). Clarke (D.L.). 1976. The Beaker network: social and economics models. In: Lanting (J.N.), Van Der Waals (J.D.), ed. Glockenbecher Symposion (Oberried, 1974). Bussum/Haarlem: Fibula-Van Dishoeck, 460-477. Clop (X.). 2002. Gestió de la matèria primera i produccions campaniformes al nord-est de la península Ibèrica. In: Pirineus i veïns al tercer mil·leni AC. De la fi del neolític a l'edat del bronze entre l'Ebre i la Garona. Institut d'Estudis Ceretans publica els treballs del XII Col·loqui Internacional d'Arqueologia de Puigcerdà, 475-488. Clop (X.). 2004. Pratiques funéraires au nord-est de la péninsule Ibérique entre 3100-1500 avant J.C. In: Besse (M.), Desideri (J.), ed. 2004. Graves and funerary rituals during the Late Neolithic and the Early Bronze Age in Europe (2700-2000 B.C.). International conference (Sion, Switzerland, October 4-7th 2001). Oxford: Archaeopress. BAR: International series S1284, 29-40. Cobourne (M.T.), Sharpe (P.T.). 2003. Tooth and jaw: molecular mechanisms of patterning in the first branchial arch. Archives of Oral Biology 48, 1-14. Constantin (C.), Blanchet (J.-C.). 1998. Le Nord de la France (Bassin Parisien). In: Guilaine (J.), ed. Atlas du Néolithique européen, 2B: l'Europe occidentale. Liège: Service de préhistoire de l’Université. Etudes et recherches archéologiques de l’Université de Liège. ERAUL 46, 585-651. Contu (E.). 1998. La cultura del vaso Campaniforme in un ipogeo di Marinaru (Sassari). In: Nicolis (F.), Mottes (E.), ed. Simbolo ed enigma: il bicchiere campaniforme e l'Italia nella preistoria europea del III millennio a.C. Catalogo della mostra (Riva del Garda, 1998). Trento: Servizio Beni Culturali, Provincia Autonoma, 286-291. Convertini (F.). 1996. Production et signification de la céramique campaniforme à la fin du 3ème millénaire av. J.C. dans le Sud et le Centre-Ouest de la France et en Suisse occidentale. Oxford: Tempus Reparatum. BAR: International series 656. Coppa (A.), Cucina (A.), Chiarelli (B.), Luna Calderon (F.), Mancinelli (D.). 1995. Dental anthropology and paleodemography of the precolumbian populations of Hispaniola from the third millennium BC to the Spanish Conquest. Human Evolution 10, 153-167. Coppa (A.), Cucina (A.), Mancinelli (D.), Vargiu (R.), Calcagno (J.M.). 1998. Dental anthropology of centralsouthern Iron Age Italy: the evidence of metric versus nonmetric traits. American Journal of Physical Anthropology 107, 371-386. Cornaggia Castiglioni (O.). 1971. La cultura di Remedello: problematica ed ergologia di un facies dell'Eneolitico padano. Milano: Museo Civico di Storia Naturale. Memorie della Sociatà Italiana di Scienze Naturali 20. Courtin (J.). 1974. Le Néolithique de la Provence. Klincksieck: Paris. Cox (G.J.), Finn (S.B.), Ast (D.B.). 1961. Effect of fluoride ingestion on the size of the cusp of Carabelli during tooth formation. Journal of Dental Research 40, 393-395. Cueni (A.). 1983. Das Dolmengrab von Aesch. Ein Beitrag zur Anthropologie und Urgeschichte. Universität Basel (Travail de diplôme non publié). Cugini (S.). 1983. Disegno e morfologia dentale. Milano: Libreria dello studente. Czarnetzki (A.). 1984. The genetic influence of Bell Beaker people in Bohemia. In: Guilaine (J.), ed. L'Age du Cuivre européen: civilisations à vases campaniformes. Paris: Editions du CNRS, 121-128. Czebreszuk (J.). 2003. Bell Beakers in the Sequence of the Cultural Changes in the South-Western Baltic Area. In: Czebreszuk (J.), Szmyt (M.), ed. The Northeast Frantier of Bell Beakers. Oxford: British Archaeological Reports. BAR: International series 1155, 21-38. 234 Bibliographie Dahlberg (A.A.). 1945. The paramolar tubercle (Bolk). American Journal of Physical Anthropology 3, 97-103. Dahlberg (A.A.). 1947. The evolutionary sequence of the protostylid. American Journal of Physical Anthropology 5, 15-25. Dahlberg (A.A.). 1950. The evolutionary significance of the protostylid. American Journal of Physical Anthropology 8, 15-24. Dahlberg (A.A.). 1956. Materials for the establishment of standards for classification of tooth characteristics, attributes and techniques in morphological studies of the dentition. University of Chicago: Zoller Laboratory of Dental Anthropology. Dahlberg (A.A.). 1961. Relationship of tooth size to cusp number and groove conformation of occlusal surface patterns of lower molar teeth. Journal of Dental Research 44, 476-479. Dahlberg (A.A.). 1963a. Analysis of the American Indian dentition. In: Brothwell (D.R.), ed. Dental anthropology. London: Pergammon Press, 149-177. Dahlberg (A.A.). 1963b. Dental evolution and culture. Human Biology 35, 237-249. Dahlberg (A.A.), Mikkelsen (O.). 1947. The shovel-shaped character in the teeth of the Pima Indians. American Journal of Physical Anthropology 5, 234-235. D'Anna (A.). 1995. La fin du Néolithique dans le Sud-Est de la France. In: Chenorkian (R.), ed. L'homme méditerranéen: mélanges offerts à Gabriel Camps, professeur émérite de l'Université de Provence. Aix-enProvence: Université de Provence, 299-333. Darlu (P.), Tassy (P.). 1993. Reconstruction phylogénétique: concepts et méthodes. Paris: Masson. Dash (J.K.), Sahoo (P.K.), Das (S.N.). 2004. Talon cusp associated with other dental anomalies: a case report. International Journal of Paediatric Dentistry 14, 295-300. Déchelette (J.). 1908. Manuel d'archéologie préhistorique, celtique et gallo-romaine, 1: Archéologie préhistorique. Paris: Picard. Del Castillo Yurrita (A.). 1928. La cultura del vaso campaniforme (Su origen y extension en Europa). Barcelona: Universidad de Barcelona. Delibes de Castro (G.), Etxeberria Gabilondo (F.). 2002. Fuego y cal en el sepulcro colectivo de "El Miradero" (Valladolid): ¿accidente, ritual o burocracia de la muerte? In: Rojo Guerra (M.), Kunst (M.), ed. Sobre el significado del fuego en los rituales funerarios del neolítico. Universidad de Valladolid: Secretariado de Publicaciones e Intercambio Editoria, 39-58 Delibes de Castro (G.), Rojo Guerra (M.). 1997. C14 y secuencia megalítica en la Lora burgalesa: acotaciones a la problemática de las dotaciones absolutas referetes a yacimientos dolménicos. In: O neolítico atlántico e as orixes do megalitismo: actas do Coloquio Internacional (Santiago de Compostela, 1-6 Abril 1996), 391-414. Delibes de Castro (G.), Rojo Guerra (M.), Sanz Mínguez (C.). 1986. Dólmenes de Sedano II. El sepulcro de corredor de Las Armillas (Moradillo de Sedano, Burgos). Madrid: Ministerio de Educación y Ciencia. Delibes de Castro (G.), Zapatero Magdaleno (P.). 1995. De lugar de habitación a sepulcro monumental: una reflexión sobre el yacimiento de la Velilla en Osorno (Palencia). Rubricatum I 1, 337-348. Demangeot (C.). (en cours). Le dénombrement de défunts dans les ensembles funéraires (problèmes théoriques, paramètres quantitatifs): application à la sépulture du Dolmen des Peirières à Villedubert, Aude, France. Université de Bordeaux I: Laboratoire des populations du Passé (Thèse de doctorat). Desideri (J.). 2001. Etude des traits épigénétiques dentaires des populations du Néolithique moyen au Bronze ancien en Suisse occidentale. Université de Genève: Département d'anthropologie et d'écologie (Travail de diplôme non publié). Desideri (J.). 2005. Les nécropoles de Barmaz I et II (Valais, Suisse): étude des traits non-métriques dentaires. Université de Genève: Département d'anthropologie et d'écologie (Rapport non publié). Desideri (J.). (à paraître). Quand l’individu témoigne…: un portrait de la société néolithique en Suisse occidentale. In: Bailly (M.), Lemercier (O.), Furestier (R.), ed. Quatrième millénaire: du Néolithique moyen au Néolithique final dans le sud-est de la France et les régions voisines. Desideri (J.), Eades (S.). 2002. La question du "peuplement" campaniforme en Suisse occidentale: état de la question et apports de l'anthropologie. Bulletins et Mémoires de la Société d’Anthropologie de Paris 14, 345362. Desideri (J.), Eades (S.). 2004. Le peuplement campaniforme en Suisse: nouveaux apports de la morphologie crânienne et dentaire. In: Besse (M.), Desideri (J.), ed. Graves and funerary rituals during the Late Neolithic and the Early Bronze Age in Europe (2700-2000 B.C.). International conference (Sion, Switzerland, October 4-7th 2001). Oxford: Archaeopress. BAR: International series S1284, 99-109. 235 Devor (E.J.), McGue (M.), Crawford (M.H.), Lin (P.M.). 1986. Transmissible and nontransmissible components of anthropometric variation in the Alexanderwohl Mennonites: II. Resolution by path analysis. American Journal of Physical Anthropology 69, 83-92. DeVoto (F.C.H.), Perrotto (B.M.). 1971. Phenotypes and genotypes of Carabelli's complex in isolated populations of Argentina highlands. Journal of Dental Research 50, 1152-1153. Devriendt (W.). 2005. Etude anthropologique de l’hypogée des Boileau: apport à la connaissance des populations du néolithique provençal. Université de la Méditerranée (Aix-Marseille II): Service d’anthropologie biologique (Thèse de doctorat non publiée). Dobes (M.). 1997. Katalog šňůrové keramiky v Čechách IX-X-Die Kataloge der Schnurkeramik in Böhmen IX-X. Praha: University Karlova. Praehistorica 22, Varia Archeologica 7, 57-112. Doro Garetto (T.), Fulcheri (E.), Crovella (S.). 1991. Manuale di antropologia dentaria. Dipartimento di Scienze, Alba (Cuneo): Museo civico Federico Eusebio. Duday (H.) 2000. De l'anthropologie de terrain à l'archéologue funéraire: le dolmen des Peirières à Villedubert (Aude, France). Notae Praehistoricae 20, 171-175. Duday (H.). 2003. De la réflexion anthropologique à l'archéologie funéraire, ou l'approche globalisante d'une sépulture collective: le Dolmen des Peirières à Villedubert (Aude, France). In: Pirineus i veïns al tercer milleni AC. De la fi del neolític a l'edat del bronze entre l'Ebre i la Garona. Institut d'Estudis Ceretans publica els treballs del XII Colloqui Internacional d'Arqueologia de Puigcerdà, 583-590. Dupanloup de Ceuninck (I.). 1999. Evaluation et synthèse des contributions de la linguistique et de la génétique à l'étude de la différentiation des populations humaines pendant la préhistoire récente. Université de Genève: Département d’anthropologie et d’écologie (Thèse de doctorat non publiée). Dürrwächter (C.), Collins (M.J.), Burger (J.), Alt (K.). 2005. Analisis de isotopos estables y analisis diagenetico de los restos oseos humanos de la Pena de Abuela y del tumulo de la Sima en la Valle de Ambrona (Soria, Espana). In: Rojo Guerra (M.A.), Kunst (M.), Garrido Pena (R.), Garcia Martinez De Lagran (I.), ed. Un Desafío a la Eternidad. Tumbas Monumentales del Valle de Ambrona (Soria, España). Junta de Castilla y León: Monografías. Arqueologia en Castilla y Leon, Memorias 14, 317-320. Dvořák (P.). 1991. Pohřebiště lidu s kulturou se zvoncovitými pohárů v Holáskách (okr. Brno-Město)-Gräberfelder der Glockenbecherkultur bei Holasky (bez. Brno-Město). Acta Musei Moraviae. Scientiae sociales 76, 41-60. Dvořák (P.), Matějíčková (A.), Peška (J.), Rakovský (I.). 1996. Die Gräberfelder der Glockenbecherkultur in Mähren, Teil 2: Katalog der Funde. Brno: P. Dvořák (Mährische archäologische Quellen). Eades (S.). 1996. Le dolmen M XII du site du Petit-Chasseur (Sion, Valais, Suisse): Paléoanthropologie de la population inhumée. Université de Genève: Département d'anthropologie et d'écologie (Travail de diplôme non publié). Eades (S.). 1997. The dental non-metric traits of the Spitalfields sample (Christ Church, London): an investigation of phenotypic similarities in a population of known family relationships. University of Bradford: Department of Archaeological Sciences (Master non publié). Eades (S.). 2003. Etude du déterminisme familial des traits non-métriques dentaires afin d'identifier des groupes d'individus apparentés en contexte funéraire archéologique. Application aux ensembles archéologiques de Kerma (Soudan), d'En Sency à Vufflens-la-Ville (Vaud, Suisse) et de Chamblandes à Pully (Vaud, Suisse). Université de Genève: Département d’anthropologie et d’écologie (Thèse de doctorat non publiée). Ebeling (C.F.), Ingervall (B.), Hegerård (B.), Lewin (T.). 1973. Secular changes in tooth in Swedish men. Acta Odontologica Scandinavica 31, 140-147. Edwards (A.W.F.). 1960. The simulation of Mendelism. Acta Genetica 10, 63-70. Efron (B.). 1979. Bootstrap methods: another look at the jackknife. Annual Statistics, 7, 1-26. Endrödi (A.). 1992. A korabronzkori Harangedény kultura telepe és temetöje Szigetszentmiklos hataraban-The settlement and cemetary of teh Bell Beaker Culture in teh district of Szigetszentmiklos. In: Havassy (P.), Selmeczi (L.), ed. Régészeti kutatasok az MO autopalya nyomvonalan, I-Archaeological researches on the line of motorway MO, I. Budapest, 83-200. Endrödi (A.). 1998. Results of settlement archeology in the Bell Beaker culture research in Hungary. In: Benz (M.), Van Willigen (S.), ed. Some new approaches to the Bell Beaker "Phenomenon": Lost Paradise...? Meeting of the Association Archéologie et Gobelets, 2 (Feldberg, Germany, April 18-20 1997). Oxford: British Archaeological Reports. BAR: International series 690, 141-160. Endrödi (A.). 2002. New data on the Late Copper Age and Early Bronze Age settlement history of Budapest-Csepel Island. Budapest: Történeti muzeum. Régiségei 36, 115-129. Endrödi (A.). 2003. Characteristic elements of the settlement structure in the early bronze age settlement at Budapest-Albertfalva. In: Kacsco (C.), ed. Bronzezeitliche kulturerscheinungen im Karpatischen raum. Die 236 Bibliographie Beziehungen zu den Benachbarten Gebieten. Ehrensymposium für Alexandru Vulpe zum 70. Geburstag (Baia Mare, 10-13 Oktober 2001). Baia Mare: Bibliotheca Marmatia 2, 151-157. Endrödi (A.), Horváth (M.). 2002. Kora bronzkori és avar kori sírok Budapest, Csepel-szigeten-Early Bronze Age and Avar graves at Budapest, Csepel-Island. In: Régészeti kutatások Magyarországon 1999 (Archaeological Investigations in Hungary 1999). Budapest, 27-45. Engelhardt (B.) 1999. Eine Kultur oder was? Archäologie in Deutschland 2, 36-38. Enoki (K.), Dahlberg (A.A.). 1958. Rotated maxillary central incisors. Orthodontic Journal of Japan 17, 157-169. Enoki (K.), Nakamura (E.). 1959. Bilateral rotation (mesiopalatal torsion) of maxillary central incisors. Journal of Dental Research 38, 204. Eogan (G.). 1976. Beaker material from Knowth, co. Meath: preliminary note In: Burgess (C.), Miket (R.), ed. Settlement and economy in the third and second millennia B.C. Conference orgonised by the Department of adult education (Newcastle upon Tyne, Jannuary 1976). Oxford: British Archaeological Reports. BAR British series 33, 251-266. Escobar (V.), Melnick (M.), Conneally (P.M.). 1976. The inheritance of bilateral rotation of central incisors. American Journal of Physical Anthropology 45, 109-115. Evans (J.A.), Chenery (C.A.), Fitzpatrick (A.P.). 2006. Bronze age childhood migration of individuals near Stonehenge, revealed by strontium and oxygen isotope tooth enamel analysis. Archaeometry 48, 309–321. Fabian Garcia (J.F.). 1995. El aspecto funerario durante el Calcolitico y los inicios de la Edad del Bronce en la Meseta Norte: el enterramiento colectivo en fosa de El Tomillar (Bercial De Zapardiel, Avila) en el marco cultural de la prehistoria reciente en el sur de la Meseta Norte espanola. Salamanca: Acta Salmanticensia (Estudios historicos y geograficos). Falconer (D.S.). 1965. The inheritance of liability to certain diseases, estimated from the incidence among relatives. Annals of Human Genetics 29, 51-76. Falconer (D.S.). 1974. Introduction à la génétique quantitative. Paris: Masson et cie. Favre (S.), Mottet (M.). 1990. Le site du Petit-Chasseur III à Sion VS: MXII, un dolmen à soubassement triangulaire du début du IIIe millénaire. Archéologie Suisse 13, 114-123. Favre (S.), Mottet (M.). 1995. Le site du Petit-Chasseur III et le dolmen MXII. In: Gallay (A.), ed. Dans les Alpes, à l'aube du métal: archéologie et bande dessinée. Catalogue d'exposition, Le Soleil des Morts: archéologie et bande dessinée (Sion, septembre 1995-janvier 1996). Sion: Musées cantonaux du Valais, 113-118. Favre (S.), Gallay (A.), Farjon (K.), Peyer (B. de). 1986. Stèles et monuments du Petit-Chasseur: un site néolithique du Valais (Suisse). Genève: Département d’anthropologie de l’Université. Ferguson (C.), Tucker (A.S.), Christensen (L.), Lau (A.), Matsuk (M.M.), Sharpe (P.T.). 1998. Activin is an essential early mesenchymal signal in tooth development that is required for patterning of the murine dentition. Genes and Development 12, 2636-2649. Fitzpatrick (A.). 2003. The Amesbury Archer. Current Archaeology 184, 146-152. Fridrich (J.). 1973. Kostrové hroby ze střední až mladší doby hradištní ve Stadicích, o. Ustí n. Labem-Körpergräber der Burgwallzeit in Stadice, Kr. Ústí n. L. (Aussig a. d. Elbe). Archeologické Rozhledy 25, 448-451. Fujiwara (T.), Nakano (S.), Sobue (S.), Ooshima (T.). 2000. Simultaneous occurrence of unusual odontodysplasia and oligodontia in the permanent dentition: report of case. International Journal of Paediatric Dentistry 10, 341-347. Furestier (R.). 2005. Les industries lithiques campaniformes du sud-est de la France. Aix-en-Provence: Université de Provence (Thèse de doctorat non publiée). Gagnière (S.). 1963. Informations archéologiques: Roquefort-les-Pins (Alpes-Maritimes). Gallia Préhistoire 6, 359361. Gallay (A.). 1975. Fouilles archéologiques du Petit-Chasseur (Sion, Valais): rapport d'activité 1973: fouille du dolmen MXI. Bolletino del Centro Camuno di Studi Preistorici 12, 103-113. Gallay (A.). 1976. The position of the Bell-Beaker civilization in the chronological sequence of Petit-Chasseur (Sion, Valais, Switzerland). In: Lanting (J.N.), Van Der Waals (J.D.), ed. Glockenbecher Symposion (Oberried, 1974). Bussum/Haarlem: Fibula-Van Dishoeck, 279-306. Gallay (A.). 1979. Le phénomène campaniforme: une nouvelle hypothèse historique. In: Menk (R.), Gallay (A.), ed. Anthropologie et archéologie: le cas des premiers âges des Métaux. International Symposium (Sils-Maria, 25-30 septembre 1978). Genève: Archives Suisses d’Anthropologie Générale 43, 231-258. Gallay (A.). 1986a. Collombey-Muraz, Barmaz I et II. In: Gallay (A.), ed. Le Valais avant l'histoire: 14 000 av. J.C.-47 ap. J.-C. Catalogue d'exposition (Sion, 23 mai-28 septembre 1986). Sion: Musées cantonaux du Valais, 184-191. 237 Gallay (A.). 1986b. Sion, Petit-Chasseur. In: Gallay (A.), ed. Le Valais avant l'histoire: 14 000 av. J.-C.-47 apr. J.-C. Catalogue d'exposition (Sion, 23 mai-28 septembre 1986). Sion: Musées cantonaux du Valais, 232-242. Gallay (A.). 1988. Le phénomène campaniforme: l'heure des remises en question. In: Beeching (A.), ed. Le Campaniforme. Arenera (Actes des rencontres Néolithique Rhône-Alpes, Lyon) 5, 6-14. Gallay (A.). 1997-1998. L'énigme campaniforme. In: Rhône-Alpes A404: l'énigmatique civilisation campaniforme. Archeologia (Dijon), Hors série, 9, 14-19. Gallay (A.). 2001. L'énigme campaniforme. In: Nicolis (F.), ed. Bell Beakers today: pottery, people, culture, symbols in prehistoric Europe: volume 1. International Colloquium (Riva del Garda, Trento, 11-16 May 1998). Trento: Servizio Beni Culturali, Provincia Autonoma di Trento, 41-58. Gallay (A.). 2004. A propos du statut épistémologique des travaux publiés sur la nécropole du Petit-Chasseur à Sion (Valais, Suisse). In: Besse (M.), Desideri (J.), ed. Graves and funerary rituals during the Late Neolithic and the Early Bronze Age in Europe (2700-2000 B.C.). International conference (Sion, Switzerland, October 47th 2001). Oxford: Archaeopress. BAR: International series S1284, 79-97. Gallay (A.), ed. 2006. Des Alpes au Léman: images de la préhistoire. Gollion: Editions Infolio. Gallay (A.), Baudais (D.). 1985. Rances, Champ-Vully Est (Vaud, Suisse). In: Première céramique, premier métal: du Néolithique à l'Âge du Bronze dans le domaine circum-alpin. Catalogue d'exposition (Lons-le-Saunier, octobre 1985-mars 1986). Lons-le-Saunier: Musée d'archéologie, 99-108. Gallay (A.), Chaix (L.). 1984. Le dolmen MXI: texte et planches, documents annexes. Lausanne: Bibliothèque historique vaudoise. Cahiers d’Archéologie Romande 31 et 32. García Barrios (A.S.). 2005. Dos singulares testimonios de cerámica simbólica en el valle medio del Duero: los restos calcolíticos de “Los Cercados” (Mucientes, Valladolid). Zephyrus 58, 245-259. García Ruiz (M.L.). 2002. Análisis de los restos óseos recuperados en el sepulcro de corredor de Las Arnillas. (Rapport non publié). Garn (S.M.), Dahlberg (A.A.), Lewis (A.B.), Kerewsky (R.S.). 1966a. Extent of sex influence on Carabelli's polymorphism. Journal of Dental Research 45, 1823. Garn (S.M.), Dahlberg (A.A.), Lewis (A.B.), Kerewesky (R.S.). 1966b. Groove pattern, cusp size and tooth size. Journal of Dental Research 45, 970. Garn (S.M.), Lewis (A.B.), Kerewsky (R.S.). 1964. Sex difference in tooth size. Journal of Dental Research 43, 306. Garn (S.M.), Lewis (A.B.), Kerewsky (R.S.). 1967. Sex difference in Tooth Shape. Journal of Dental Research 46, 1470. Garn (S.M.), Lewis (A.B.), Vicinus (J.H.). 1962. Third molar agenesis and reduction in the number of other teeth. Journal of Dental Research 41, 717. Garn (S.M.), Osborne (R.H.), Alvesalo (L.), Horowitz (S.L.). 1980. Maternal and gestational influence on deciduous and permanent tooth size. Journal of Dental Research 42, 142-143. Garralda (M.D.). 1979. Les populations du Néolithique et du Bronze I dans la péninsule Ibérique et les îles Baléares. In: Menk (R.), Gallay (A.), ed. Anthropologie et archéologie: le cas des premiers âges des Métaux. International Symposium (Sils-Maria, 25-30 septembre 1978). Genève: Archives Suisses d’Anthropologie Générale 43, 211-222. Garrido-Pena (R.). 2000. El Campaniforme en la Meseta Central de la Península Ibérica (c. 2500-2000 A.C.). Oxford: Archaeopress. BAR: International series 892. Garrido-Pena (R). 2005. The Beaker Labyrinth: Brief history of an intellectual challenge-El Laberinto Campaniforme: Breve historia de un reto intelectual. In: Rojo Guerra (M.A.), Garrido Pena (R.), Garcia Martinez De Lagran (I.), ed. El Campaniforme en la Peninsula Ibérica y su contexto europeo-Bell Beakers in the Iberian Peninsula and their european context. Valladolid: Universidad de Valladolid. Arte y Arqueologia 21, 29-60. Garrido-Pena (R.), Rojo Guerra (M.), Garcia Martinez De Lagran (I.). 2005. Bell Beakers In Central Iberia (the Inner Meseta)-El Campaniforme en la Meseta central de la Peninsula Ibérica. In: Rojo Guerra (M.A.), Garrido Pena (R.), Garcia Martinez De Lagran (I.), ed. El Campaniforme en la Peninsula Ibérica y su contexto europeo-Bell Beakers in the Iberian Peninsula and their european context. Valladolid: Universidad de Valladolid. Arte y Arqueologia 21, 411-456. Gelbert (A.). 2003. Traditions céramiques et emprunts techniques: étude ethnoarchéologique dans les haute et moyenne vallées du fleuve Sénégal (+ CD-ROM). Paris: Editions de la Maison des sciences de l'hommeEpistèmes. (Collection Référentiels). Gerhardt (K.). 1953. Die Glockenbecherleute in Mittel-und Westdeutschland: ein Beitrag zur Paläoanthropologie Eurafrikas. Stuttgart: E. Schweizerbart'sche Verlagsbuchhandlung. 238 Bibliographie Gerhardt (K.). 1976. Anthropotypologie der Glockenbecherleute in ihren Ausschwärmelandschaften. In: Lanting (J.N.), Van Der Waals (J.D.), ed. Glockenbecher Symposion (Oberried, 1974). Bussum/Haarlem: Fibula-Van Dishoeck, 147-166. Gersbach (E.). 1966-1967. Zur Herkunft und Zeitstellung der einfachen Dolmen vom Typus Aesch-Schwörstadt. Annuaire de la Société Suisse de Préhistoire et d’Archéologie 53, 15-28. Goldstein (M.S.). 1931. The cusp in the mandibular teeth of the Eskimo. American Journal of Physical Anthropology 15, 215-235. Gonzalez Martin (A.), Campo Martin (M.). 2002. Estudio antropologico de los restos oseos recuperados de la necropolis del Valle de las Higueras (Huecas, Toledo). (Rapport non publié). Goose (D.H.), Lee (G.T.R.). 1971. The mode of inheritance of Carabelli's trait. Human Biology 43, 64-69. Grainger (R.M.), Paynter (K.J.), Shaw (J.H.). 1959. Differences in the morphology and size of the teeth of a cariessusceptible and a caries-resistant strain of rats. Journal of Dental Research 38, 105-120. Gravlee (C.C.), Bernard (H.R.), Leonard (W.R.). 2003a. Boas’s changes in bodily form: the immigrants study, cranial plasticity, and Boas’s physical anthropology. American Anthropologist 105, 326-332. Gravlee (C.C.), Bernard (H.R.), Leonard (W.R.). 2003b. Heredity, environment, and cranial form: a reanalysis of Boas’s immigrant data. American Anthropologist 105, 125-138. Greene (D.L.). 1982. Discrete and dental variations and biological distances of Nubian populations. American Journal of Physical Anthropology 58, 75-79. Gregory (W.K.). 1916. Studies on the evolution of the primates. Bulletin of the American Museum of Natural History 35, 239-357. Grüneberg (H.). 1952. Genetical studies on the skeleton of the mouse. IV. Quasi-continuous variations. Journal of Genetics 51, 95-114. Guagliardo (M.F.). 1982. Tooth crown size differences between age groups: a possible new indicator of stress in skeletal samples. American Journal of Physical Anthropology 58, 383-389. Guilaine (J.). 1967. La civilisation du vase campaniforme dans les Pyrénées françaises. Carcassonne: Imprimerie Gabelle. Guilaine (J.). 1998a. Atlas du Néolithique européen ; 2A, 2B: L'Europe occidentale. Liège: Service de préhistoire de l’Université. Etudes et recherches archéologiques de l’Université de Liège. ERAUL 46. Guilaine (J.). 1998b. Au temps des dolmens: mégalithes et vie quotidienne en France méditerranéenne il y a 5000 ans. Toulouse: Privat. Guilaine (J.). 1998c. Impressions de Riva del Garda: Bell Beakers Today: pottery, people, culture, symbols in third millenium Europe. Bulletin de la Société Préhistorique Française 95, 418-419. Guilaine (J.), ed. 2007. Le Chalcolithique et la construction des inégalités, 1: le continent européen. Paris: Editions Errance (Collection des Hespérides). Guilaine (J.), Blanchet (J.-C.), L'Helgouach (J.), Petrequin (P.), Roussot-Laroque (J.). 1988. Le Chalcolithique en France. In: L'età del Rame in Europa. Congresso internazionale (Viareggio, 15-18 ottobre 1987). Firenze: All'Insegna del Giglio. Rassegna di Archeologia 7, 211-263. Guilaine (J.), Vaquer (J.), Coularou (J.), Treinen-Claustre (F.), et al. 1989. Ornaisons-Médor: archéologie et écologie d'un site de l'Âge du Cuivre, de l'Âge du Bronze final et de l'Antiquité tardive. Toulouse: Centre d'anthropologie des sociétés rurales, Carcassonne: Archéologie en Terre d'Aude. Guilaine (J.), Claustre (F.), Lemercier (O.), Sabatier (P.). 2001. Campaniformes et environnement culturel en France Méditerranéenne. In: Nicolis (F.), ed. Bell Beakers today: pottery, people, culture, symbols in prehistoric Europe: vol. 1. International Colloquium (Riva del Garda, Trento, 11-16 May 1998). Trento: Servizio Beni Culturali, Provincia Autonoma di Trento, 229-275. Haeussler (A.M.), Irish (J.D.), Morris (D.H.), Turner (C.G.). 1989. Morphological and metrical comparison of San and Central Sotho dentitions from southern Africa. American Journal of Physical Anthropology 78, 115-122. Hájek (L.). 1957. Knoflíky středoevropské skupiny kultury zvoncovitych pohárů-Die Knöpfe der mitteleuropäischen Glockenbecherkultur. Památky Archeologické 48, 389-424. Hájek (L.). 1966. Die älteste Phase des Glockenbecherkultur in Böhmen und Mären. Památky Archeologické 57, 210-241. Hájek (L.). 1968. Die Glockenbecherkultur in Böhmen-Kultura zvoncovitých pohárů v Čechách. Praha: Archeologický ústav Čsav. Archeologické studijní materiály 5. Halffman (C.M.), Scott (G.R.), Pedersen (P.O.). 1992. Palatine torus in the Greenlandic Norse. American Journal of Physical Anthropology 88, 145-161. Hammer (O.), Harper (D.). 2005. Paleontological Data Analysis. Oxford: Blackwell publishing. 239 Hanihara (K.). 1961. Criteria for classification of crown characters of the human deciduous dentition. Journal of the Anthropological Society of Nippon 69, 27-45 (in Japanese with English summary). Hanihara (K.). 1963. Crown characters of the deciduous dentition of the Japanese-American hybrids. In: Brothwell (D.R.), ed. Dental anthropology. London: Pergammon Press, 105-124. Hanihara (K.). 1967. Racial characteristics in the dentition. Journal of Dental Research 46, 923-926. Hanihara (K.). 1976. Statistical and comparative studies of the Australian Aboriginal dentition. Tokyo: University of Tokyo Press. Bulletin 11, 28-37. Hanihara (K.), Tamada (M.), Tanaka (T.). 1970. Quantitative analysis of the hypocone in the human upper molar. Journal of the Anthropological Society of Nippon 78, 200-207. Harila (V.). 2004. The effect of preterm birth on the development of the dentition. University of Oulu: Acta Universitatis Ouluensis Medica (Dissertation non publiée). Harris (E.F.). 1980. Sex differences in lingual marginal ridging on the human maxillary central incisor. American Journal of Physical Anthropology 52, 541-548. Harris (E.F.), Bailit (H.L.). 1980. The metaconule: a morphologic and familial analysis of a molar cusp in humans. American Journal of Physical Anthropology 53, 349-358. Harris (E.F.), Dinh (D.P.). 2006. Intercusp relationships of the permanent maxillary first and second molars in American Whites. American Journal of Physical Anthropology 130, 514-528. Harrison (R.J.). 1974. Origins of the Bell Beaker cultures. Antiquity 48, 99-109. Harrison (R.J.). 1980. The Beaker folk: Copper Age archeology in Western Europe. London: Thames and Hudson. Harrison (R.J.). 1986. L'Age du Cuivre: la civilisation du vase campaniforme. Paris: Editions Errance (Collection des Hespérides). Harrison (R.J.), Mederos Martin (A.). 2001. Bell Beakers and social complexity in Central Spain. In: Nicolis (F.), ed. Bell Beakers today: pottery, people, culture, symbols in prehistoric Europe: volume 1. International Colloquium (Riva del Garda, Trento, 11-16 May 1998). Trento: Servizio Beni Culturali, Provincia Autonoma di Trento, 111-124. Hauser (G.), De Stefano (G.F.). 1989. Epigenetic variants of the human skull. Stuttgart: Schweizerbart. Havel (J.). 1978. Pohřební ritus kultury zvoncovitých pohárů v Čechách a na Moravě. The Burial Rite of the Bell Beaker Culture in Bohemia and Moravia. Praha: University Karlova. Praehistorica 7, Varia Archeologica 1, 91-117. Havel (J.). 1981. Nové nálezy únětické kultury v Praze 8-Čimicích-Neue Funde der Úněticer Kultur in Prag 8Čimice. Archaeologica Pragensia 2, 71-81. Havel (J.). 1982. Dva hroby kultury zvoncovitých pohárů z Prahy, Archaeologica Pragensia 3, 5-12. Hébras (C.). 1965. Le Dolmen E 136, du groupe de Monpalais, commune de Taizé (Deux-Sèvres). Bulletin de la Société Préhistorique Française 62, 139-158. Heiss (M.H.), Kolar (L.), Luksha (L.), Johnson (C.). 1998. Biology of the human dentition. Chicago: University of Illinois (document online: www. uic.edu/classes/orla/orla312). Hénon (P.), Vérot Bourrely (A.). 1998. Habitats néolithiques, protohistoriques et occupations historiques du site de "Derrière-le-Château", à Géovreissiat et Montréal-la-Cluse-Ain-France. In: Frere-Sautot (M.-C.), ed. Paléométallurgie des cuivres. Actes du colloque (Bourg-en-Bresse et Beaune, 17-18 octobre 1997). Montagnac: Monique Mergoil, 207-249. Heyd (V.), 2004. Die Siedlungsspuren der Glockenbecherkultur unter dem Kapellplatz von Altötting. In: Heyd (V.), Husty (L.), Kreiner (L.), ed. 2004. Siedlungen der Glockenbecherkultur in Süddeutschland und Mitteleuropa. Arbeiten zur Archäologie Süddeutschlands 17, 121-146. Heyd (V.), Husty (L.), Kreiner (L.). 2004. Siedlungen der Glockenbecherkultur in Süddeutschland und Mitteleuropa. Arbeiten zur Archäologie Süddeutschlands 17. Higa (T.), Hanihara (T.), Sunakawa (H.), Ishida (H.). 2003. Dental variation of Ryukyu Islanders: A comparative study among Ryukyu, Ainu and other Asian population. American Journal of Human Biology 15, 127-143. Hillson (S.). 1986. Teeth. Cambridge: Cambridge University Press. Hillson (S.). 1996. Dental anthropology. Cambridge: Cambridge University Press. Hnizdova (I.), Simunek (J.). 1955. Hrob se šňůrovou keramikou v Blšanech u Loun-Sépulture a céramique cordée a Blšany près de Louny. Archeologické Rozhledy 5, 577-582. Holloway (R.L.). 2002. Head to head with Boas: did he err on the plasticity of head form? Proceedings of the National Academy of Sciences of the United States of America 99, 23, 14622-14623. Holloway (R.L.). 2004. La forme du crâne, un vrai casse-tête… La Recherche 377, 58-59. 240 Bibliographie Honegger (M.). 1994-1995. Données récentes sur les nécropoles du Néolithique moyen I et du Bronze ancien de Barmaz I (Collombey-Muraz, Valais). In: Aspects culturels et religieux: témoignages et évolution de la préhistoire à l'an mil. Actes du 7e colloque international sur les Alpes dans l'Antiquité (Châtillon, Vallée d'Aoste, 11-13 mars 1994). Aoste: Bulletin d'études préhistoriques et archéologiques alpines 5/6, numéro spécial, 53-66. Honegger (M.). 1995. Barmaz I, Collombey-Muraz. In: Gallay (A.), ed. Dans les Alpes, à l'aube du métal: archéologie et bande dessinée. Catalogue d'exposition, Le Soleil des Morts: archéologie et bande dessinée (Sion, septembre 1995-janvier 1996). Sion: Musées cantonaux du Valais, 85-90. Honegger (M.), Desideri (J.). 2003. Archéologie et anthropologie de la nécropole du Néolithique moyen de Barmaz (Valais, Suisse). In: Chambon (P.), Leclerc (J.), ed. Les pratiques funéraires néolithiques avant 3500 av. J.-C. en France et dans les régions limitrophes. Table ronde SPF (Saint-Germain-en-Laye, 15-17 juin 2001). Paris: Société préhistorique française. Mémoires de la Société Préhistorique Française 33, 219-230. Horan (G.), Ramirez-Solis (R.), Featherstone (M.), Wolgemuth (D.), Bradley (A.) Behringer (R.). 1995. Compound mutants for the paralogous Hoxa-4, Hoxb-4, and Hoxd-4 genes show more complete homeotic transformations and a dose-dependent increase in the number of vertebrae transformed. Genes and Development 9, 1667-1677. Horváth (M.), Andras-Szilas (G.), Endrödi (A.), Horváth (M.A.). 2004. Megelösö feltaras Dunakeszi-SzékesdülönPreliminary excavation at Dunakeszi-Székesdülön. In: Öskoros Kutatok II. Összejövetelének konferenciakötete. ΜΩΜΣ ΙΙ, 209-218. Hrala (J.), Jiráň (L.), Moucha (V.). 1994. Studies of the Bronze Age in Bohemia: the last twenty five years. In: Fridrich, (J.), ed. 25 Years of archaeological research in Bohemia. Prague: Institute of Archaeology. Památky archeologické, supplementum, 55-72. Hrdlickà (A.). 1920. Shovel-shaped teeth. American Journal of Physical Anthropology 3, 429-465. Hrdlickà (A.). 1921. Further studies of tooth morphology. American Journal of Physical Anthropology 4, 141-176. Hrdlickà (A.). 1924. New data on the teeth of early man and certain fossil European apes. American Journal of Physical Anthropology 7, 109-132. Hsu (J.W.), Tsai (P.L.), Hsiao (T.H.), Chang (H.P.), Lin (L.M.), Liu (K.M.), Yu (H.S.), Ferguson (D.). 1999. Ethnic dental analysis of shovel and Carabelli’s traits in a Chinese population. Australian Dental Journal 44, 40-45. Huguenin (C.), Moinat (P.). 1998. La nécropole néolithique de Pully-Chamblandes: l'avènement de l'archéologie funéraire. In: Bertholet (D.), Feihl (O.), Huguenin (C.), ed. Autour de Chillon: archéologie et restauration au début du siècle. Catalogue d'exposition (Lausanne, 4 septembre-27 décembre 1998). Lausanne: Musée cantonal d'archéologie et d'histoire, 83-88. Hunter (J.P.), Jernvall (J.). 1995. The hypocone as key innovation in mammalian evolution. Proceedings of the National Academy of Sciences of the United States of America 92, 10718-10722. Hutsy (L.). 2004. Ueberlegungen zum Siedlungswesen der Glockenbecherkultur. In: Heyd (V.), Husty (L.), Kreiner (L.), ed. 2004. Siedlungen der Glockenbecherkultur in Süddeutschland und Mitteleuropa. Arbeiten zur Archäologie Süddeutschlands 17, 147-154. Irish (J.D.). 1998. Ancestral dental traits in recent Sub-Saharan Africans and the origins of modern humans. Journal of Human Evolution 34, 81-98. Irish (J.D.), Morris (D.H.). 1996. Canine mesial ridge (Bushman Canine): Dental trait definition. American Journal of Physical Anthropology 99, 357-359. Irish (J.D.), Turner (C.G.). 1990. West African dental affinities of Late Pleistocene Nubians peopling the EurafricanSouth Asian triangle II. Homo 41, 42-53. Jernvall (J.), Thesleff (I.). 2000. Reiterative signalling and patterning during mammalian tooth morphogenesis. Mechanics of Development 92, 19-29. Jeunesse (C.). 1998. Compte-rendu de Benz (M.) et van Willigen (S.), ed. (1998). Bulletin de la Société Préhistorique Française 95, 415-417. Jimenez Sanz (P.J.), Alcolea González (J.J.). 2000. Excavaciones arqueológicas en la cueva del Destete Valdepeñas de la Sierra, Guadalajara: cuestiones preliminares. Actas del primer Simposio de Arqueología de Guadalajara (Sigüenza, 4-7 octubre 2000), 293-308. Johnson (A.L.), Lovell (N.). 1995. Dental morphological evidence for biological continuity between the A-Group and C-Group periods in lower Nubia. International Journal of Osteoarchaeology 5, 368-376. Jørgensen (K.D.). 1956. The deciduous dentition: a descriptive and comparative anatomical study. Acta Odontologica Scandinavica 16, 1-202. 241 Joshi (M.R.). 1975. Carabelli's trait on maxillary second deciduous molars in Hindus. Archives of Oral Biology 20, 699-700. Kalicz-Schreiber (R.). 1981. Möglichkeiten zur feineren Gliederung der Nagyrév-Kultur in Budapest. In: Die Frühbronzezeit im Karpatenbecken und in Nachbargebieten. International Symposium (Budapest-Velem, 1977). Budapest: Archäologischer Institut der UAW. Mitteilungen des archäologischen Instituts der ungarischen Akademie der Wissenschaften. Beiheft 2, 81-86. Kalicz-Schreiber (R.), Kalicz (N.), 2001. Were the Bell Beakers as social indicators of the early Bronze Age of Budapest ? In: Nicolis (F.), ed. Bell Beakers today: pottery, people, culture, symbols in prehistoric Europe: volume 2. International Colloquium (Riva del Garda, Trento, 11-16 May 1998). Trento: Servizio Beni Culturali, Provincia Autonoma di Trento, 439-458. Kanasawa (E.), Sekikawa (M.), Ozaki (T.). 1990. A quantitative investigation of irregular cuspules in human maxillary permanent molars. American Journal of Physical Anthropology 83, 173-180. Kassai (Y.), Munne (P.), Hotta (Y.), Penttilä (E.), Kavanagh (K.), Ohbayashi (N.), Takada (S.), Thesleff (I.), Jernvall (J.), Itoh (N.). 2005. Regulation of mammalian tooth cusp patterning by ectodin. Science 309, 20672070. Kau (C.H.), Hunter (M.L.), Lewis (B.), Knox (J.). 2003. Tooth agenesis: a report of missing molars in two generations. International Journal of Paediatric Dentistry 13, 342-347. Kangas (A.T.), Evans (A.R.), Thesleff (I.), Jernvall (J.). 2004. Nonindependence of mammalian dental characters. Nature 432, 211-214. Kaul (V.), Prakash (S.). 1981. Morphological features of Jat dentition. American Journal of Physical Anthropology 54, 123-127. Kaul (S.S.), Sharma (K.), Sharma (J.C.), Corrucini (R.S.). 1985. Non-metric variants of the permanent crown in human monozygous and dizygous twins. In: Reddy (V.R.), ed. Dental anthropology: application and methods. New-Dehli: Inter-India publications, 187-193. Keene (H.J.). 1982. The morphogenetic triangle: a new conceptual tool for application to problems in dental morphogenesis. American Journal of Physical Anthropology 59, 281-287. Keränen (S.V.E.), Åberg (T.), Kettunen (P.), Thesleff (I.), Jernvall (J.). 1998. Association of developmental regulatory genes with the development of different molar tooth shapes in two species of rodents. Development Genes and Evolution 208, 477-486. Kieser (J.A.). 1990. Human Adult Odontometrics. Cambridge University Press, Cambridge. Kieser (J.A.), Groeneveld (H.T.). 1988. Fluctuating odontometric asymmetry in an urban South African black population. Journal of Dental Research 67, 1200-1205. Kieser (J.A.), Preston (C.B.). 1981. The Dentition of the Lengua Indians of Paraguay. American Journal of Physical Anthropology 55, 485. Kirveskari (P.). 1978. Racial traits in the dentition of living Skolt Lapps. In: Butler (P.M.), Joysey (K.A.), ed. Development, function and evolution of teeth. London: Academic Press, 59-68. Knor (A.). 1955. Druhé únětické pohřebiště v Brodcích nad Jizerou-Deuxième nécropole ouniétitzienne de Brodce, arrt Mladá Boleslav. Archeologické Rozhledy 7, 582-588. Kobyliansky (E.) 1983. Changes in cephalic morphology of Israelis due to migration. Journal of Human Evolution 12, 779-786. Kohn (L.A.P). 1991. The role of genetics in craniofacial morphology and growth. Annual Review of Anthropology 20, 261-278. Kolakowski (D.), Bailit (H.L.). 1981. Segregation analyses of tooth size in a Melanesian population. Annals of Human Biology 8, 255-262. Kolakowski (D.), Harris (E.F.). 1981. A differential environmental effect on human anterior tooth size. American Journal of Physical Anthropology 54, 377-381. Kolakowski (D.), Harris (E.F.), Bailit (H.L.). 1980. Complex segregation analysis of Carabelli's trait in a Melanesian population. American Journal of Physical Anthropology 53, 301-308. Koutecký (D.). 1966. Archeologický výzkum ve Vikleticích v letech 1961-1964-Archäologische Forschungen in Vikletice in den Jahren 1961-1964. Archeologické Rozhledy 18, 121-129. Koutecký (D.). 1968. Šňůrové pohřebiště v Polákách-Das schnurkeramische Gräberfeld in Poláky. Archeologické Rozhledy 20, 289-291. Koutecký (D.), Smrz (Z.). 1991. Pohřebiště bylanské kultury v Polákách, okr. Chomutov I. Památky Archeologické 82, 166-230. 242 Bibliographie Kovařík (J.). 1980. Hroby se šňůrovou keramikou a únětické pohřebiště v Praze-Čakovicích-Gräber der schnurkeramischen Kultur und das Gräberfeld der Úněticer Kultur in Prag 9-Čakovice. Archeologica Pragensia 1, 55-99. Kovařík (J.). 1981. Praha 5-Malá Ohrada. Praha: Pražský sborník historický XIII, 183. Kramar (C.). 1977. Les restes humains néolithiques du premier dolmen (MVI) du Petit-Chasseur à Sion (VS). Genève: Archives Suisses d’Anthropologie Générale 41, 37-42. Kramar (C.). 1982. Nécropole de Corseaux-sur-Vevey: étude anthropologique et description archéologique. Université de Genève: Département d’anthropologie et d’écologie (Thèse de doctorat non publiée). Kramar (C.), Sauter (M.-R.), Weidmann (D.). 1978. La nécropole néolithique de Corseaux-sur-Vevey. Archéologie Suisse 1, 51-54. Kraus (B.S.). 1951. Carabelli's anomaly of the maxillary molar teeth. Observations on Mexican and Papago Indians and an interpretation of the inheritance. American Journal of Human Genetics 3, 348-355. Kraus (B.S.). 1959. Occurrence of the Carabelli trait in Southwestern ethnic groups. American Journal of Human Genetics 3, 348-355. Kraus (B.S.), Furr (M.L.). 1953. Lower first premolar: a definition and classification of discrete morphologic traits. Part 1. Journal of Dental Research 32, 554-564. Krueger (B.J.). 1962. Influence of boron, fluorine, and molybdenum on dental morphology of the rat molar. Journal of Dental Research 41, 215. Krueger (B.J.). 1966. Interaction of fluoride and molybdenum on dental morphology in the rat. Journal of Dental Research 45, 714-725. Krutova (M.). 2003. Bell Beaker and Unetice burial rites. Continuity and change in funerary practices at the beginning of Bronze Age. In: Czebreszuk (J.), Szmyt (M.), ed. The Northeast Frontier of Bell Beakers. Poznan: Proceedings of the symposium held at the Adam Mickiewicz University, 209-214. Kytlicová (O.). 1956a. Neolitické a mohylové sídliště v Kněževsi-Stations néolithiques et des tumuli à Kněževes. Archeologické Rozhledy 8, 777-782. Kytlicová (O.). 1956b. Pohřebiště kultury zvoncovitých pohárů v Kněževsi-Nécropole de la civilisation à vases campaniformes à Kněževes, arrt Prague-ouest. Archeologické Rozhledy 8, 328-336. Kytlicová (O.). 1957. Hrob se šňůrovou keramikou z Kněževse-Das Grab mit Schnurkeramik in Kněževes (Böhmen). Archeologické Rozhledy 9, 571-573. Kytlicová (O.). 1960. Eneolitické pohřebiště v Brandýsku-Das äneolitische Gräberfeld in der Gemeinde Brandýsek. Památky Archeologické 61, 442-474. Kytlicová (O.). 1965. Neolitické a eneolitické osídlení v Mochově-Die neolithische und äneolithische Siedlung in Mochov. Archeologické Rozhledy 17, 178-182. Larson (M.J.), Bader (R.S.). 1976. Maternal Effects in Odontometric Traits of House Mice. American Midland Naturalist 96, 21-35. Lasker (G.W.). 1945. Observations on the teeth of Chinese born and reared in China and America. American Journal of Physical Anthropology 3, 129-150. Lanting (J.N.). 1973. Laat-neolithicum en Vroege Bronstijd in Netherland en N.W.-Duitsland: continue ontwikkelingen. Palaeohistoria, 15, 216-317. Lanting (J.N.), Van Der Waals (J.D.). 1976. Beaker culture relations in the lower Rhine Basin. In: Lanting (J.N.), Van Der Waals (J.D.), ed. Glockenbecher Symposion (Oberried, 1974). Bussum/Haarlem: Fibula-Van Dishoeck, 1-80. Lee (G.T.R.), Goose (D.H.). 1982. Heritability of dental occlusal variables in a family study in Liverpool, UK. Archives of Oral Biology 27, 987-989. Leisner (G.), Leisner (V.). 1943. Die Megalithgräber der Iberischen Halbinsel, I: der Süden. Berlin: W. de Gruyter. Lemercier (O.). 1997-1998. Le Campaniforme dans la moyenne et basse vallée du Rhône. In: Rhône-Alpes A404: l'énigmatique civilisation campaniforme. Archeologia (Dijon), Hors série, 9, 30-35. Lemercier (O.). 2002. Le Campaniforme dans le sud-est de la France. De l’Archéologie à l’Histoire du troisième millénaire avant notre ère. Aix-en-Provence: Université de Provence (Thèse de doctorat non publiée). Lemercier (O.). 2003. La périodisation du Campaniforme dans le Midi. Jean Guilaine avait raison. In: ADAL, ed. Temps et Espaces culturels du 6° au 2° millénaire en France du sud. Rencontres Méridionales de Préhistoire Récente, IVe session, (Nîmes, 28-29 octobre 2000), 151-160. Lemercier (O.). 2004. Les Campaniformes dans le sud-est de la France. Lattes: Monographies d’Archéologie Méditerranéenne 18. 243 L'Helgouach (J.). 1998. L'Ouest de la France. In: Guilaine (J.), ed. Atlas du Néolithique européen, 2B: l'Europe occidentale. Liège: Service de préhistoire de l’Université. Etudes et recherches archéologiques de l’Université de Liège. ERAUL 46, 653-688. Lička (M.). 1967. Kostrový hrob kultury volutové z Trmic (o. Ústí n. L.)-Ein Körpergrab der bandkeramischen Kultur aus Trmice, Krs. Ústí nad Labem. Archeologické Rozhledy 19, 87-90. Line (S.R.P.). 2001. Molecular Morphogenetic Fields in the Development of Human Dentition. Journal of Theorical Biology 211, 67-75. Liversage (D.). 2003. Bell Beaker pottery in Denmark: Its Typology and Internal Chronology. In: Czebreszuk (J.), Szmyt (M.), ed. The Northeast Frontier of Bell Beakers. Poznan: Proceedings of the symposium held at the Adam Mickiewicz University, 39-50. Loh (H.S.). 1998. Root morphology of the maxillary first premolar in Singaporeans. Australian Dental Journal 43, 399-402. Louwe Kooijmans (L.). 1974. The Rhine/Meuse delta: four studies on its prehistoric occupation and Holocen geology. Leiden: E.J. Brill. Ludwig (F.S.). 1957. The mandibular second premolars: Morphologic variation and inheritance. Journal of Dental Research 36, 263-273. Lukacs (J.R.). 1983a. Dental anthropology and the origins of Iron-Age populations from northern Pakistan. Homo 34, 1-15. Lukacs (J.R.). 1983b. Human Dental Remains From Early Neolithic Levels at Mehrgarh, Baluchistan. Current Anthropology 24, 390-392. Lundström (A.). 1963. Tooth morphology as a basis for distinguishing monozygotic and dizygotic twins. American Journal of Human Genetics 15, 34-43. Machnik (J.). 1984. Frühbronzezeitliche Kulturen in Kleinpolen. In: Tasic (N.), ed. Kulturen der Frühbronzezeit das Karpatenbeckens und Nordbalkans. Beograd: Balkanoloski Institut Sanu. Balcano-Pannonica 22, 341-365. MacKusick (V.A.). 1990. Mendelian inheritance in man: a catalog of human genes and genetic disorders. Baltimore: Johns Hopkins University Press. Mahieu (E.). 1987. L'hypogée des Boileau (Sarrians, Vaucluse). (Rapport de fouille non publié). Mahieu (E.). 1989. L'hypogée des Boileau. Marseille: Etudes et Prospectives Archéologiques. Mahieu (E.). 1992a. Les sépultures campaniformes en France. In: Barge-Mahieu (H.), ed. Le Campaniforme dans le Midi de la France: origine et identité culturelle. Exposition (Orgon, 6 juillet-28 septembre 1992). Marseille: Etudes et Prospectives Archéologiques, 74-79. Mahieu (E.). 1992b. Premiers apports de l'hypogée des Boileau à l'étude des sépultures collectives du Sud-Est de la France. In: Mahieu (E.), ed. Anthropologie préhistorique: résultats et tendances. Colloque (Sarrians, 2-4 septembre 1989). Sarrians: Etudes et Prospectives Archéologiques, 75-81. Mahieu (E.), Devriendt (W.), Barge (H.), Lalys (L.), Adalian (P.), Yann Ardagna (Y.), Signoli (M.). 2004. L'hypogée des Boileau (Vaucluse, France): Etude fonctionnelle et anthropologique. In: Section 9. Le Néolithique au Proche Orient et en Europe-The Neolithic in the Near East and Europe. Section 10: L'âge du cuivre au Proche Orient et en Europe-The Copper Age in the Near East and Europe. Actes du XIVème Congrès UISPP Université de Liège (University of Liège, Belgium, September 2-8th 2001). Oxford: Archaeopress. BAR: International series S1303, 375-383. Makarowicz (P.). 2003. Northern and southern Bell Beaker in Poland. In: Czebreszuk (J.), Szmyt (M.), ed. The Northeast Frantier of Bell Beakers. Oxford: British Archaeological Reports. BAR: International series 1155, 137-154. Manabe (Y.), Rokatunda (A.), Kitagawa (Y.). 1992. Nonmetric tooth crown traits in the Ami tribe, Taiwan aborigenes: comparisons with other East Asian populations. Human Biology 64, 717-726. Marieb (E.N.). 1999. Anatomie et physiologie humaines. Bruxelles: De Boeck Université. Marseiller (E.). 1984. Les dents humaines: morphologie. Paris: Gauthier-Villars. Marti Oliver (B.), Juan-Cabanilles (J.). 1998. L’Espagne méditerranéenne: pays valencien et région de Murcie. In: Guilaine (J.), ed. Atlas du Néolithique européen, 2B: l'Europe occidentale. Liège: Service de préhistoire de l’Université. Etudes et recherches archéologiques de l’Université de Liège. ERAUL 46, 825-870. Martin Carbajo (A.M.), Misiego Tejeda (J.C.), Perez Rodriguez (F.J.), Fernandez Gimenez (J.M.), Sanz Garcia (F.J.), Marcos Contreras (G.J.). 1993. Documento funerario del bronce medio en la submeseta norte: Carrelasvegas (Santilla de Campos, Palencia). Boletín del Seminario de Estudios de Arte y Arqueología 59, 69-88. 244 Bibliographie Martin Colliga (A.). 1998. Le nord-est de la Péninsule Ibérique (et les Baléares). In: Guilaine (J.), ed. Atlas du Néolithique européen, 2B: l'Europe occidentale. Liège: Service de préhistoire de l’Université. Etudes et recherches archéologiques de l’Université de Liège. ERAUL 46, 763-824. Martin Montes (M.A.), Perez Rodriguez-Aragon (F.). 1997. Un nuevo asentamiento neolítico al aire libre en la Meseta Norte: La Cañadilla de Torre de Peñafiel (Valladolid). Boletín del Seminario de Estudios de Arte y Arqueología 63, 31-48. Martin Socas (D.), Camalich Massieu (M.D.), Gonzalez Quintero (P.). 1998. L’Andalousie. In: Guilaine (J.), ed. Atlas du Néolithique européen, 2B: l'Europe occidentale. Liège: Service de préhistoire de l’Université. Etudes et recherches archéologiques de l’Université de Liège. ERAUL 46, 871-933. Martin Valls (R.), Delibes de Castro (G.). 1989. La cultura del vaso campaniforme en las campinas merdidionales del Duero: El enterramiento de Fuente-Olmedo (Valladolid). Junta de Castilla y Leon: Monografias del museo arqueologico de Valladolid 1. Masters (D.H.), Hoskine (S.W.). 1964. Projection of cervical enamel into molar furcations. Journal of Periodontology 35, 49-53. Mayhall (J.T.). 1992. Techniques for the study of dental morphology. In: Saunders (S.R.), Katzenberg (M.A.), ed. Skeletal biology of past peoples: research methods. New York: Wiley-Liss, 59-78. Mayhall (J.T.), Alvesalo (L.). 1992. Dental morphology of 45, XO human females: molar cusp area, volume, shape and linear measurements. Archives of Oral Biology 37, 1039-1043. Mayhall (J.T.), Saunders (S.R.), Belier (P.L.). 1982. The dental morphology of American Whites: a reappraisal. In: Kurten, (B.), ed. Teeth: form, function, evolution. New York: Columbia University Press, 245-258. Mays (S.). 1998. The Archaeology of Human Bones. United Kindom: Routledge. Mays (S.). 2005. Talon cusp in a primary lateral incisor from a medieval child. International Journal of Paediatric Dentistry 15, 67-72. Medunová (A.), Ondraček (J.). 1969. Birituální pohrebiste lidu s kulturou zvoncovitych poháru u Lechovice, okr. Znojmo-Das birituelle Gräberfeld der Glockenbecherkultur bei Lechovice, Kr. Znojmo. Archeologické Rozhledy 21, 437-445. Menk (R.), 1979. Le phénomène campaniforme: structures biologiques et intégration historique. In: Menk (R.), Gallay (A.), ed. Anthropologie et archéologie: le cas des premiers âges des Métaux. International Symposium (Sils-Maria, 25-30 septembre 1978). Genève: Archives Suisses d’Anthropologie Générale 43, 259-284. Menk (R.). 1981. Anthropologie du Néolithique européen: analyse multivariée et essai de synthèse. Université de Genève: Département d'anthropologie et d'écologie (Thèse de doctorat non publiée). Meredith (H.V.), Hixon (E.H.). 1954. Frequency, size and bilateralism of Carabelli’s tubercle. Journal of Dental Research 33, 435-440. Mezzena (F.). 1985. La vallée d'Aoste dans la préhistoire et la protohistoire. In: Archéologie en Vallée d'Aoste: du Néolithique à la chute de l'Empire romain, 3500 av. J.-C.-Vème siècle apr. J.-C. Catalogue d'exposition, Château Sarriod de la Tour (Saint-Pierre, 22 août 1981). Aosta: Assessorato del Turismo, Urbanisme et Biens culturels, 15-60. Mezzena (F.). 1995. Area megalitica di Aosta. In: Gambari (M.V.), Poggiani Keller (R.), Mezzena (F.), ed. Guide archeologiche: Preistoria e Protostoria in Italia, 8: Lombardia occidentale, Piemonte e Valle d'Aosta. International congress of prehistoric and protohistoric sciences, 13 (Forlì, 8-14 septembre 1996). Forlì: A.B.A.C.O, 102-125. Miletich (I.), Sharpe (P.T.). 2003. Normal and abnormal dental development. Human Molecular Genetics 12, 69-73. Millán (A.), Arribas (J.G.). 1994. La ceramica: estudio tecnologico. In: Blasco (C.), ed. El horizonte campaniforme de la region de Madrid en el centenario de Ciempozuelos. Madrid: Univerisdad autonoma, 117-126. Mitsiadis (E.A.). 2001. Bases moléculaires du développement dentaire. In: Piette (E.), Goldberg (M.), ed. La dent normale et pathologique. Bruxelles: De Boeck Université, 19-38. Mizoguchi (Y.). 1985. Shoveling: A Statistical Analysis of its Morphology. Tokyo: University of Tokyo Press Bulletin 26, 1-176. Mizoguchi (Y.). 1988. Degree of bilateral asymmetry of nonmetric tooth crown characters quantified by the tetrachoric correlation method. Bulletin of the National Science Museum 14, 29-49. Mizoguchi (Y.). 1989. Genetic variability of left-right asymmetries and mirror imagery in nonmetric tooth crown characters. Bulletin of the National Science Museum 15, 49-61. Modat (J.-F.). 2003. Biométrie, morphologie et pathologie du matériel dentaire isolé du dolmen chalcolithique des des Peyraoutes (Roquefort-les-Pins, Alpes-Maritimes). Université de la Méditerranée (Aix-Marseille II): Laboratoire d’anthropologie, secteur Nord (Diplôme d’études avancées non publié). 245 Moinat (P.). 1994. Cistes néolithiques et incinération du Bronze final à Pully VD-Chamblandes. Annuaire de la Société suisse de préhistoire et d'archéologie 77, 123-126. Moinat (P.), Gallay (A.). 1998. Les tombes de type Chamblandes et l'origine du mégalithisme alpin. Archéologie Suisse 21, 2-12. Moinat (P.), Simon (C.). 1985. La nécropole néolithique de Chamblandes (Pully, VD). In: Première céramique, premier métal: du Néolithique à l'Age du Bronze dans le domaine circum-alpin. Catalogue d'exposition (Lons-le-Saunier, octobre 1985-mars 1986). Lons-le-Saunier: Musée d'archéologie, 109-113. Moinat (P.), Simon (C.). 1986. Nécropole de Chamblandes-Pully: nouvelles observations. Annuaire de la Société suisse de préhistoire et d'archéologie 69, 39-53. Moinat (P.), Stöckli (W.E.). 1995. Croyances et rites funéraires. In: Stöckli (W.E.), Niffeler (U.), Gross-Klee (E.), ed. Néolithique. Bâle: Société suisse de préhistoire et d'archéologie. SPM: La Suisse du Paléolithique à l'aube du Moyen-Age 2, 231-258. Møller (I.J.). 1967. Influence of Microelements on the Morphology of the Teeth. Journal of Dental Research 46, 933-937. Montelius (O.). 1900. Die Chronologie der ältesten Bronzezeit in Norddeutschland und Skandinavien. Archiv für Anthropologie (Braunschweig) 25, 1-40. Moorrees (C.F.A.). 1951. The dentition as a criterion of race with special reference to the Aleut. Journal of Dental Research 30, 815-821. Morin (A.). 1998. Les niveaux B1 et B2 de l'abri sous Bloc n°1. In: Bintz (P.), ed. Les sites archéologiques de l'Aulp du Seuil à Saint-Bernard-du-Touvet (Chartreuse, Isère: l'abri sous Bloc n°1). Grenoble: Université Joseph Fourier. (Rapport scientifique non publié). Morris (D.H.). 1970. On deflecting wrinkles and the Dryopithecus pattern in human mandibular molars. American Journal of Physical Anthropology 32, 97-104. Morris (D.H.). 1975. Bushman maxillary canine polymorphism. South African Journal of Science 71, 333-335. Morton (N.E.), MacLean (C.J.). 1974. Analysis of family resemblance III. Complex segregation analysis of quantitative traits. American Journal of Human Genetics 26, 489-503 Morton (N.E.), Yee (S.), Lew (R.). 1971. Complex segregation analysis. American Journal of Human Genetics 23, 602-611. Moskona (D.), Vainder (M.), Hershkovitz (I.), Koyliansky (E.). 1997. Dental morphological (non-metric) traits in human isolates: South Sinai Bedouin tribes. Homo 48, 227-284. Moucha (V.). 1958. Beitrag zur Frage der zeitlichen Einordnung äneolithischer Gürtelplatten. Archeologické Rozhledy 10, 62-78. Moucha (V.). 1963. Die Periodisierung der Úněticer Kultur in Böhmen. Sborník Československé společnosti archeologické při ČSAV 3, 9-60. Moucha (V.). 1966. Nécropole à Mochov (Bohême). Investigations archéologiques en Tchécoslovaquie 107. Moucha (V.). 1978. VI:14 Kultura se zvoncovitymi poháry, In: Pleiner (R.), Rybová (A.), ed. Pravěké dějiny Čech. Praha, 300-312. Moucha (V.). 1992. Die Schnurkeramik und die Glockenbecherkultur in Böhmen. In: Buchvaldek (M.), Strahm (C.), ed. Die kontinentaleuropäischen Gruppen der Kultur mit Schnurkeramik. Schnurkeramik Symposium (Praha, 1990). Praha: University Karlova. Praehistorica 19, 81-87. Müller (A.). 1998. Geschlechtsspezifische Totenlage und geschlechtsspezifische Beigaben bei der BöhmischMährischen Gruppe bzw. Ostgruppe der Glockenbecherkultur. In: Benz (M.), Van Willigen (S.), ed. Some new approaches to the Bell Beaker "Phenomenon": Lost Paradise...? Meeting of the Association Archéologie et Gobelets, 2 (Feldberg, Germany, April 18-20 1997). Oxford: British Archaeological Reports. BAR: International series 690, 121-127. Müller (J.), Van Willigen (S.). 2001. New radiocarbon evidence for european Bell Beakers and the consequences for the diffusion of the Bell Beaker Phenomenon. In: Nicolis (F.), ed. Bell Beakers today: pottery, people, culture, symbols in prehistoric Europe: volume 1. International Colloquium (Riva del Garda, Trento, 11-16 May 1998). Trento: Servizio Beni Culturali, Provincia Autonoma di Trento, 59-80. Muška (J.) 1981. The settlement and cemetery sites of the Bell Beaker culture at Radovesice by Bílina. Xe Congrès International des sciences préhistoriques et protohistoriques (Prague-Brno), 51. Naef (A.). 1901. La nécropole néolithique de Chamblandes (canton de Vaud). L'anthropologie 12, 269-276. Naugler (C.T.), Ludman (M.D.). 1996. Fluctuating asymmetry and disorders of developmental origin. American Journal of Medical Genetics 66, 15-20. Nelson (C.T.). 1938. The teeth of the Indians of Pecos Pueblo. American Journal of Physical Anthropology 23, 261293. 246 Bibliographie Neugebauer (J.-W.), Gattringer (A.). 1982. Die Kremser Schnellstrasse S 33. Fundberichte aus Österreich 21, 63-96. Neugebauer (J.-W.), Neugebauer-Maresch (C.). 2001. Bell Beaker Culture in Austria. In: Nicolis (F.), ed. Bell Beakers today: pottery, people, culture, symbols in prehistoric Europe: volume 2. International Colloquium (Riva del Garda, Trento, 11-16 May 1998). Trento: Servizio Beni Culturali, Provincia Autonoma di Trento, 429-438. Neugebauer-Maresch (C.). 1994. Endneolithikum. In: Neugebauer (J.-W.), ed. Bronzezeit in Ostösterreich. St. Pölten-Wien: Verlag Niederösterreich. Pressehaus. Wissenschaftliche Schriftenreihe Niederösterreich 98101, 23-48. Neustupný (E.). 1984. The Bell Beaker Culture in East Central Europe. In: Guilaine (J.), ed. L'Âge du Cuivre européen: civilisations à vases campaniformes. Paris: Editions du CNRS, 107-119. Neustupný (E.), Smrž (Z.). 1989. Čachovice: pohřebistě kultury se śňůrovou keramikou a zvoncovitých pohárůČachovice: a Corded Ware and Bell Beaker Cemetary. Památky Archeologické 80, 282-383. Nichol (C.R.). 1989. Complex segregation analysis of dental morphologic variants. American Journal of Physical Anthropology 78, 37-59. Nichol (C.R.). 1990. Dental genetics and biological relationships of the Pima Indians of Arizona. Arizona State University: Department of anthropology (PhD dissertation non publiée). Nichol (C.R.), Turner II (C.G.). 1986. Intra- and interobserver concordance in classifying dental morphology. American Journal of Physical Anthropology 69, 299-315. Nichol (C.R.), Turner II (C.G.), Dahlberg, (A.A.). 1984. Variation in the convexity of the human maxillary incisor labial surface. American Journal of Physical Anthropology 63, 361-370. Nicklisch (N.), Uepermann (A.), Alt (K.). 2005. Estudios de los restos humanos neoliticos de la tumba de la Sima, Mino de Medinaceli. In: Rojo Guerra (M.A.), Kunst (M.), Garrido Pena (R.), Garcia Martinez De Lagran (I.), ed. Un Desafío a la Eternidad. Tumbas Monumentales del Valle de Ambrona (Soria, España). Junta de Castilla y León: Monografías. Arqueologia en Castilla y Leon, Memorias 14, 323-339. Nicolis (F.), Mottes (E.). 1998. Simbolo ed enigma: il bicchiere campaniforme e l'Italia nella preistoria europea del III millennio a.C. Catalogo della mostra (Riva del Garda, 1998). Trento: Servizio Beni Culturali, Provincia Autonoma. Nieminen (P.), Arte (S.), Pirinen (S.), Peltonen (L.), Thesleff (I.). 1995. Gene defect in hypodontia: exclusion of MSX1 and MSX2 as candidate genes. Human Genetics 96, 305-308. Nosek (S.). 1964. Studia i materialy do badan nad neolitem Malopolski. Wroclaw-Warszawa-Kraków. Noss (J.F.), Scott (G.R.), Potter (R.H.Y), Dahlberg (A.A.). 1983. Fluctuating asymmetry in molar dimensions and discrete morphological traits in Pima Indians. American Journal of Physical Anthropology 61, 437-445. Osborn (J.W.). 1978. Morphogenetic gradients: field versus clones. In: Butler (P.M.), Joysey (K.A.), ed. Development, function and evolution of teeth. London: Academic Press, 171-201. Ossenberg (N.S.). 1976. Within and between race distances in population studies based on discrete traits of human skull American Journal of Physical Anthropology 45, 701-715. Otto (K.-H.). 1950. Neue Gräber des Glockenbecherfriedhofs von Schafstädt. Kr. Merseburg. Jahresschrift für mitteldeutsche Vorgeschichte 34, 55-80. Palliardi (J.). 1919. Beiträge zur Kenntnis der Glockenbecherkultur. Wiener Prähistorische Zeitschrift 4, 41-56. Palomino (H.), Chakraborty (R.), Rothhammer (F.). 1977. Dental morphology and population diversity. Human Biology 49, 61-70. Palosaari (H.). 2003. Matrix metalloproteinases (MMPs) and their specific tissue inhibitors (TIMPs) in mature human odontoblasts and pulp tissue. The regulation of expressions of fibrillar collagens, MMPs and TIMPs by growth factors, transforming growth factor-β1 (TGF-β1) and bone morphogenetic protein-2 (BMP-2). Acta Universitatis Ouluensis D 739. University of Oulu: Institute of dentistery (Dissertation non publiée). Parras (Z.). 2004. The biological affinities of the Eastern Mediterranean in the Chalcolithic and Bronze Age. A regional dental non-metric approach. Oxford: British Archaeological Reports. BAR: International series 1305. Pedersen (P.O.). 1949. The East Greenland Eskimo dentition: numerical variations and anatomy. Copenhagen: Meddelelser om Grønland 3, 1-244. Pemberton, (T.J.), Gee (J.), Patel (P.I.). 2006. Gene discovery for dental anomalies. Journal of the American Dental Association 137, 743-752. Perzigian (A.J.). 1976. The dentition of the Indian Knoll skeletal population: odontometrics and cusp number. American Journal of Physical Anthropology 44, 113-122. Perzigian (A.J.). 1984. Human odontometric variation: an evolutionary and taxonomic perspective. Anthropologie 22, 193-198. 247 Piera (S.). 2000. Etude des caractères discrets et de l’état sanitaire bucco-dentaire dans les cairns de Fontenay-lemarmion (Calvados): un recrutement familial des inhumés dans les tombes à couloir du Néolithique moyen? Université de Paris I: Laboratoire de protohistoire européenne (Diplôme d’études avancées non publié). Pirinen (S.), Arte (S.), Apajalahti (S.). 1996. Palatal displacement of canine is genetic and related to congenital absence of teeth. Journal of Dental Research 75, 1742-1746. Pirinen (S.), Kentala (A.), Nieminen (P.), Varilo (T.), Thesleff (I.), Arte (S.). 2001. Recessively inherited lower incisor hypodontia. Journal of Medical Genetics 38, 551-556. Pispa (J.), Thesleff (I.). 2003. Mechanisms of ectodermal organogenesis. Developmental Biology 262, 195-205. Pleinerová (I.). 1960. Únětické pohřebiště a osada v Blšanech u Loun-Gräberfeld und Siedlung der Aunjetitzer Kultur von Blšany bei Laun. Památky Archeologické 51, 488-526. Pleinerová (I.). 1965. Výzkum osady z doby stěhování národů a z doby slovanské v Březně u Loun-Erforschung einer Siedlung aus der Völkerwanderungs- und Slawenzeit in Březno bei Louny. Archeologické Rozhledy 17, 495-500. Pleinerová (I.). 1990. Hrob kultury zvoncovitých pohárů ze Selibic-Ein Grab der Glockenbecherkultur bei Selibice. Archeologické Rozhledy 42, 207-209. Plesl (E.). 1959. Nálezy únětické kultury na mohylníku v Mladé Boleslavi-Čejetičkách-Funde der Aunjetitzer Kultur auf dem Hügelgräberfeld in Mladá Boleslav-Čejetičky. Památky Archeologické 50, 34-53. Polly (P.D.). 2006. Genetics, development, and palaeontology interlock. Heredity 96, 206-207. Popelka (M.). 1995. Štípaná industrie z Malého Března, okr. Most. In: Blažek (J.), Meduna (P.), ed. Archeologické výzkumy v severozápadních Čechách v letech 1983-1992, 97-110. Portin (P.), Alvesalo (L.). 1974. The inheritance of shovel shape in maxillary central incisors. American Journal of Physical Anthropology 41, 59-62. Potter (R.H.). 1990. Twin Half-sibs: A Research Design for Genetic Epidemiology of Common Dental Disorders. Journal of Dental Research 69, 1527-1530. Potter (R.H.), Nance (W.E.). 1976. A twin study of dental dimension. I. Discordance, asymmetry, and mirror imagery. American Journal of Physical Anthropology 44, 391-396. Price (T.D.), Grupe (G.), Schröter (P.). 1998. Migration and mobility in the Bell Beaker period in Central Europe. Antiquity 72, 405-411. Price (T.D.), Knipper (C.), Grupe (G.), Smrcka (V.). 2004. Strontium Isotopes and Prehistoric Human Migration: The Bell Beaker Period in Central Europe. European Journal of Archaeology 7, 9-40. Prieto Martinez (M.P.). 1998. Forma, estilo y contexto en la cultura material de la Edad del Bronce gallega: cerá campaniforme y cerámica no decorada. Universidad de Santiago de Compostella: Facultad de Geografia e Historia (Thèse de doctorat non publiée). Qiu (M.), Bulfone (A.), Ghattas (I.), Meneses (J.J.), Christensen (L.), Sharpe (P.T.), Presley (R.), Pedersen (R.A.), Rubenstein (J.L.). 1997. Role of Dlx-1 and -2 in proximodistal patterning of the branchial arches: mutations alter morphogenesis of proximal skeletal elements derived from the first and second arches. Developmental Biology 185, 165-184 Rada (I.). 1981. Únětické pohřebiště v Chotěbudicích, okr. Louny-The Únětician culture cemetery at Chotěbudice, the Louny distr. (viz Chochol J.). Archeologické Rozhledy 33, 621-634. Rataj (J.). 1954. Konstrukce únětických hrobek v Brodcích nad Jizerou. Construction des tombeaux ouniétitziennee de Brodce-sur-Jizera, Bohéme. Archeologické Rozhledy 6, 154-158. Rataj (J.). 1955. Stavby únětických hrobů v Brodcích nad Jizerou. Tombeaux ouniétitziens de Brodce-sur-Jizera. Archeologické Rozhledy 7, 292-293. Rataj (J.). 1958. Únětické pohřebiště na Hrušově-Das Aunjetitzer Gräberfeld in Hrušov (Bez. Ml. Boleslav), Böhmen. Archeologické Rozhledy 10, 305-312. Rehman (F.), Robinson (V.J.), Shennan (S.J.). 1992. A neutron activation study of Bell Beakers and associated pottery from Czechoslovakia und Hungary. Památky Archeologické 83, 97-211. Reid (C.), Reenen (J.R. von), Groeneveld (H.T.). 1991. Tooth size and the Carabelli trait. American Journal of Physical Anthropology 84, 427-432. Reichart (P.), Tantiniran (D.). 1975. Dens evaginatus in the Thai. An evaluation of fifty-one cases. Oral Surgery, Oral Medecine, Oral Pathology 39, 615-621. Reinecke (P.). 1900. Zur jüngeren Steinzeit in West- und Süddeutschland. Westdeutsche Zeitschrift für Geschichte und Kunst (Trier) 19, 209-270. Riquet (R.), Guilaine (J.), Coffyn (A.). 1963. Les Campaniformes français. Gallia Préhistoire 6, 63-128. 248 Bibliographie Roger (J.-M.). 1992. La station de Maupas à Calvisson (Gard) et le Campaniforme en Languedoc oriental. In: Barge-Mahieu (H.), ed. Le Campaniforme dans le Midi de la France: origine et identité culturelle. Exposition (Orgon, 6 juillet-28 septembre 1992). Marseille: Etudes et Prospectives Archéologiques, 55-58. Rojo Guerra (M.A.), Garrido Pena (R.), Garcia Martinez De Lagran (I.). 2005a. El Campaniforme en la Peninsula Ibérica y su contexto europeo-Bell Beakers in the Iberian Peninsula and their european context. Arte y Arqueologia 21. Valladolid: Universidad de Valladolid. Rojo Guerra (M.A.), Kunst (M.), Palomino (A.L.). 2002. El fuego como procedimiento de clausura en tres tumbas monumentales de la Submeseta Norte. In: Rojo Guerra (M.A.), Kunst (M.), ed. Sobre el Significado del Fuego en los Rituales Funerarios del Neolítico. Valladolid: Studia Archaeologica 91, 21-38. Rojo Guerra (M.A.), Kunst (M.), Garrido Pena (R.), Garcia Martinez De Lagran (I.). 2005b. Un Desafío a la Eternidad. Tumbas Monumentales del Valle de Ambrona (Soria, España). Junta de Castilla y León: Monografías. Arqueologia en Castilla y Leon, Memorias 14. Rojo Guerra (M.A.), Moran Dauchez (G.), Flores Fernandez (R.), Latorre Navalpotro (J.L.). 2003. El Pasato Escrito en el Paisaje. Guía de Recursos Culturales y Medioambientales del Valle de Ambrona. Soria: Ochoa Impresores. Rothhammer (F.), Lasserre (E.), Blanco (R.), Covarrubias (E.), Dixon (M.). 1968. Microevolution in Chilean populations. VI. Shovel shape, mesial-palatal version and other dental traits in Pewneche Indians. Zeitschrift für Morphologie und Anthropologie 60, 162-169. Roudil (J.-L.), Bazile (F.), Soulier (M.). 1974. L'habitat campaniforme de Saint-Côme-et-Maruéjols (Gard). Gallia Préhistoire 17, 181-217. Roussot-Larroque (J.). 1990. Paradigmes perdus, paradigmes retrouvés... Le Campaniforme atlantique et les sociétés du Néolithique final de l'Ouest. Revue archéologique de l'Ouest, supplément, 2, 189-204. Ruch (J.V.). 2001. Développement dentaire normal. In: Piette (E.), Goldberg (M.), ed. La dent normale et pathologique. Bruxelles: De Boeck Université, 5-18. Rusó (A.). 1985. Prosetice, okr. Teplice. Výzkumy v Čechách 1982-1983, 149. Salanova (L.). 1997. La céramique décorée du site campanifome de Derrière-le-Château (Géovreissiat, Ain). (Rapport non publié) Salanova (L.). 2000. La question du campaniforme en France et dans les îles anglo-normandes: productions, chronologie et rôles d'un standard céramique. Paris: Editions du Comité des travaux historiques et scientifiques (C.T.H.S.). Salanova (L.). 2002. Fabrication et circulation des céramiques campaniformes. In: Guilaine (J.), ed. Matériaux, productions, circulations du Néolithique à l'âge du Bronze. Paris: Editions Errance. (Collection des Hespérides), 151-166. Salanova (L.). 2003. Les sépultures mégalithiques et le phénomène campaniforme. In: Gonçalves (V.S.), ed. Muita gente, poucas antas ? Origens, espaçios e contextos do Megalitismo. Actes du IIe colloque international sur le Mégalithisme, (Reguengos de Monsaraz, 3-7 maio 2000). Trabalhos de Arqueologia 25, 385-393. Salanova (L.). 2005. Los origenes del campaniforme: descomponer, analizar, cartografiar-The origins of the Bell Beaker phenomenon: breakdown, analysis, mapping. In: Rojo Guerra (M.A.), Garrido Pena (R.), Garcia Martinez De Lagran (I.), ed. El Campaniforme en la Peninsula Ibérica y su contexto europeo-Bell Beakers in the Iberian Peninsula and their european context. Valladolid: Universidad de Valladolid. Arte y Arqueologia 21, 7-27. Salanova (L.), Ducreux (F.), ed. 2005. L’habitat campaniforme de La Noue à Saint-Marcel (Saône-et-Loire). Eléments de définition du groupe Bourguignon-Jurassien. Gallia Préhistoire 47, 33-146. Salanova (L.), Sohn (M.). 2007. Mobilier funéraire et modes de différenciation des morts à la fin du Néolithique en Europe occidentale. In: Baray (L.), Brun (P.), Testart (A.), ed. Pratiques funéraires et sociétés. Nouvelles approches en archéologie et en anthropologie sociale. Dijon: Editions Universitaires de Dijon. Art, Archéologie et Patrimoine, 77-90. Salazar-Ciudad (I.), Jernvall (J.). 2002. A gene network model accounting for development and evolution of mammalian teeth. Proceedings of the National Academy of Sciences 99, 8116-8120. Sangmeister (E.). 1963. Exposé sur la civilisation du vase campaniforme. In: Les civilisations atlantiques du Néolithique à l'âge du Fer. Colloque atlantique 1 (Brest, 11 septembre 1961). Rennes: Laboratoire d'anthropologie préhistorique, 25-55. Sangmeister (E.), Schubart (H.). 1981. Zambujal. Die Grabungen1964 bis 1973. Maienz. Sarazin (F.). 1910. Das steinzeitliche Dolmengrab bei Aesch unweit Basel. Verhandlungen der Naturforschenden Gesellschaft in Basel 21, 266-291. Sarti (L.). 1998. Il Campaniforme di Neto-Via Verga (Sesto Fiorentino, Firenze). Riviste di scienze preistoriche 48, 367-398. 249 Sarti (L.), Martini (F.). 1998. Il tumulo di via Bruschi a Sesto Fiorentino. In: Nicolis (F.), Mottes (E.), ed. Simbolo ed enigma: il bicchiere campaniforme e l'Italia nella preistoria europea del III millennio a.C. Catalogo della mostra (Riva del Garda, 1998). Trento: Servizio Beni Culturali, Provincia Autonoma, 168-173. Sarti (L.), Martini (F.). 2001. Strategie insediative de Campaniforme in Italia centrale. In: Nicolis (F.), ed. Bell Beakers today: pottery, people, culture, symbols in prehistoric Europe: volume 1. International Colloquium (Riva del Garda, Trento, 11-16 May 1998). Trento: Servizio Beni Culturali, Provincia Autonoma di Trento, 187-198. Saunders (S.R.), Mayhall (J.T.). 1982. Developmental patterns of human dental morphological traits. Archives of Oral Biology 27, 45-49. Saunders (S.R.), Mayhall (J.T.). 1986. Fluctuating asymmetry of dental morphological traits: New interpretations. Human Biology 54, 789-799. Sauter (M.-R.). 1949. Les nécropoles de la Barmaz sur Collombey (Valais). Fouilles de 1948. La Suisse Primitive 13, 10-14. Sauter (M.-R.). 1950. Collombey (Valais). Les dernières fouilles de la nécropole de la Barmaz, 1950. La Suisse Primitive 14, 45-47. Sauter (M.-R.). 1951. Collombey (Valais). La Barmaz II. Fouilles de 1951. La Suisse Primitive 15, 52-54. Sauter (M.-R.). 1979. Essais de mise en relation entre les groupes néolithiques de Barmaz I et II (Collombey-Muraz VS) et de Chamblandes (Pully, VD). Genève: Archives Suisses d’Anthropologie Générale 43, 165-181. Sauter (M.-R.). 1985. Notes posthumes sur les cimetières néolithiques de Barmaz I et II (Collombey-Muraz, Valais, Suisse). Aoste: Bulletin d’Etudes Préhistoriques Alpines 17, 19-32. Sauzade (G.). 1983. Les sépultures du Vaucluse du Néolithique à l'Age du Bronze. Paris: Editions du Laboratoire de paléontologie humaine et de préhistoire. Etudes quaternaires, mémoires 6. Sauzade (G.). 1987. Dolmen I des Cudières à Jouques (Bouches-du-Rhône). (Rapport de fouille non publié). Sauzade (G.). 1988. Dolmen I des Cudières à Jouques (Bouches-du-Rhône). (Rapport de fouille non publié). Schäffler (A.), Menche (N.). 2004. Anatomie, physiologie, biologie. Paris: Maloine. Schenk (A.). 1901. Les sépultures et les populations préhistoriques de Chamblandes. Revue Historique Vaudoise 9/8, 241-250. Schenk (A.). 1904. Les squelettes préhistoriques de Chamblandes (Suisse). Revue de l'Ecole d'anthropologie de Paris 14, 335-378. Scheuer (L.), Black (S.). 2000. Developmental Juvenile Osteology. London: Elsevier Academic Press. Schliz (A.). 1910. Die vorgeschichtlichen Schädeltypen der deutschen Länder in ihrer Beziehung zu den einzelnen Kulturkreisen der Urgeschichte. Archiv für Anthropologie 9, 202-251. Schmidt (H.). 1913. H. Schmidt, Estudios acerca de los principios de la Edad de los Metales en España. Madrid: Comisión de InvestigacionesPaleontológicas y Prehistóricas, Memorias 8. Schulze (C.). 1987. Anomalien and Missbildungen des menschlissen Zähne. Berlin: Quintessenz. Sciulli (P.W.). 1977. A descriptive and comparative study of the deciduous dentition of Prehistoric Ohio Valley Amerindians. American Journal of Physical Anthropology 47, 71-80. Sciulli (P.W.). 1998. Evolution of the dentition in prehistoric Ohio valley Native Americans: II. Morphology of the deciduous dentition. American Journal of Physical Anthropology 106, 189-205. Scott (G.R.). 1973. Dental morphology: a genetic study of American White families and variation in living Southwest Indians. Arizona State University: Department of anthropology (PhD dissertation non publiée). Scott (G.R.). 1975. Association between nonmetrical tooth crown characteristics. American Journal of Physical Anthropology 42, 328. Scott (G.R.). 1977. Classification, sex dimorphism, association and populational variation of the canine distal accessory ridge. Human Biology 49, 453-469. Scott (G.R.). 1978. The relationship between Carabelli’s trait and the protostylid. Journal of Dental Research 57, 570. Scott (G.R.). 1979. Association between the hypocone and Carabelli’s trait of the maxillary molars. Journal of Dental Research 58, 1403-1404. Scott (G.R.). 1980. Population variation of Carabelli's trait. Human Biology 52, 63-78. Scott (G.R.), Alexandersen (V.). 1992. Dental morphological variation among medieval Greenlanders, Icelanders, and Norwegians. In: Smith (P.), Tchernov (E.), ed. Structure, Function and Evolution. London: Freund Publishing House Ltd, 467-490. 250 Bibliographie Scott (G.R.), Dahlberg (A.A.). 1982. Microdifferentiation in tooth crown morphology among Indians of the American Southwest. In: Kurten (B.), ed. Teeth: form, function, evolution. New York: Columbia University Press, 259-291. Scott (G.R.), Potter (R.H.). 1984. An analysis of tooth crown morphology in American white twins. Anthropologie 22, 223-231. Scott (G.R.), Turner II (C.G.). 1997. The anthropology of human modern teeth, Dental morphology and its variation in recent human populations. Cambridge: University Press. Scott (G.R.), Potter (R.H.Y.), Noss (J.N.), Dahlberg (A.A.), Dahlberg (T.). 1983. The dental morphology of Pima Indians. American Journal of Physical Anthropology 61, 13-31. Searle (A.G.). 1954a. Genetical studies on the skeleton of the mouse. IX. Causes of skeletal variation within pure lines. Journal of Genetics 52, 68-102. Searle (A.G.). 1954b. Genetical studies on the skeleton of the mouse. XI. The influence of diet on variation within pure lines. Journal of Genetics 52, 413-424. Sharma (K.) 1992. Dental morphology and odontometry of twins and the heritability of dental variation. In: Lukacs (J.R.), ed. Culture, ecology and dental anthropology. Journal of Human Evolution Special Issue no 2. Delhi: Kamla-Raj Enterprises, 49-60. Sharpe (P.T.). 1995. Homeobox genes and orofacial development. Connective Tissue Research 32, 17-25. Sharpe (P.T.). 2001. Neural crest and tooth morphogenesis. Advances in Dental Research 15, 4-7. Shaw (J.C.M.). 1927. Cusp development on second lower molar. American Journal of Physical Anthropology 11, 97-190. Shennan (S.J.). 1976. Bell Beakers and their context in Central Europe. In: Lanting (J.N.), Van Der Waals (J.D.), ed. Glockenbecher Symposion (Oberried, 1974). Bussum/Haarlem: Fibula-Van Dishoeck, 231-239. Simon (C.), Kaufmann (B.). 1998. Anthropologie: remarques préliminaires. In: Hochuli (S.), Niffeler (U.), Rychner (V.), ed. Age du Bronze. Bâle: Société suisse de préhistoire et d'archéologie. SPM: La Suisse du Paléolithique à l'aube du Moyen-Age 3, 347-354. Sjøvold (T.). 1977. Non-metrical divergence between skeletal populations. Ossa: International Journal of Skeletal Research 4, supplément, 1, 1-133. Sladek (V.), Berner (M.), Sailer (R.). 2006a. Mobility in Central European Late Eneolithic and Early Bronze Age: Femoral Cross-Sectional Geometry. American Journal of Physical Anthropology 130, 320–332. Sladek (V.), Berner (M.), Sailer (R.). 2006b. Mobility in Central European Late Eneolithic and Early Bronze Age: tibial cross-sectional geometry. Journal of Archaeological Science 33, 470-482. Smejtek (L.), Vojtěchovská (I.). 1997. Velké Přílepy 1994-1995. Předběžné výsledky a strategie výzkumu zanikající lokality-Velké Přílepy 1994-1995. Vorläufige Ergebnisse und Forschungsstrategien einer untergehend Lokalität. Archeologické Rozhledy 49, 9-18. Smith (B.H.), Garn (S.M.), Cole (P.E.). 1982. Problems of sampling and inference in the study of fluctuating asymmetry. American Journal of Physical Anthropology 58, 281-289. Snyder (R.G.). 1960. Mesial marging ridging of incisor labial surfaces. Journal of Dental Research 39, 361-364. Sofaer (J.A.). Chung (C.S.), Niswander (J.D.), Runck (D.W.). 1971. Developmental interaction, size and agenesis among permanent maxillary incisors. Human Biology 43, 36. Sofaer (J.A.), MacLean (C.J.), Bailit (H.L.). 1972a. Heredity and morphological variation in early and late developing teeth of the same morphological class. Archives of Oral Biology 17, 811-816. Sofaer (J.A.), Niswander (J.D.), MacLean (C.J.). 1972b. Population studies on Southwest Indian tribes. V: Tooth morphology as an indicator of biological distance. American Journal of Physical Anthropology 37, 357-366. Šolle (M.). 1949. Nové hroby ze Sulejovic u Lovosic-Nouvelles tombes à Sulejovice, Bohême. Archeologické Rozhledy 1, 163-164. Šolle (M.). 1966. Stará Kouřim a projevy velkomoravské hmotné kultury v Čechách. Monumenta Archaeologica 15. Sparks (C.S.), Jantz (R.L.). 2002. A reassessment of human cranial plasticity: Boas revisited. Proceedings of the National Academy of Sciences of the United States of America 99, 23, 14636-14639. Sparks (C.S.), Jantz (R.L.). 2003. Changing times, changing faces: Franz Boas’s immigrant study in modern perspective. American Anthropologist 105, 333-337. Sperber (G.H.). 2004. The Genetics of Odontogenesis: Implications in Dental Anthropology and Palæo-Odontology. Dental Anthropology 17, 1-7. Stahlhofen (H.). 1986. Drei Gräber der späten Glockenbecherkultur von Gross Börnecke, Kr. Stassfurt. Ausgrabungen und Funde (Berlin) 31, 156-160. 251 Staley (R.N.), Green (L.J.). 1974. Types of tooth cusp occurrence asymmetry in human monozygotic and dizygotic twins. American Journal of Physical Anthropology 40, 187-195. Staley (R.N.), Vance (J.P.), Loucks (C.D.). 1998. Brief communication: Type II tooth cusp occurrence asymmetry in a human monozygotic twin pair. American Journal of Physical Anthropology 105, 93-95. Stephen (L.X.G.), Basel (D.), Beighton (P.H.). 2002. Developmental absence of the premolar teeth: dental management. International Journal of Paediatric Dentistry 12, 219–222. Stevens (A.), Lowe (J.). 1996. Anatomie pathologique générale et spéciale. Bruxelles: De Boeck Université. Strahm (C.). 1995. Das Glockenbecher-Phänomen: ein Seminar. Freiburg Im Brisgau: Institut für Ur- und Frühgeschichte der Universitat. Freiburger archäologische Studien 2. Strahm (C.). 1998. Il bicchiere campaniforme: fenomeno e cultura. In: Nicolis (F.), Mottes (E.), ed. Simbolo ed enigma: il bicchiere campaniforme e l'Italia nella preistoria europea del III millennio a.C. Catalogo della mostra (Riva del Garda, 1998). Trento: Servizio Beni Culturali, Provincia Autonoma, 21-44. Strahm (C.). 2004. Le phénomène campaniforme et les composantes autochtones non campaniformes. Bulletin de la Société Préhistorique Française 101, 201-206. Strahl (E.). 1990. Das Endneolithikum im Elb-Weser-Dreieck, Teil 1: Text, Teil 2: Katalog, Teil 3: Tafeln und Karten. Hildesheim: A. Lax. Veröffentlichungen der urgeschichtlichen Sammlungen des Landesmuseums zu Hannover 36. Suarez (B.K.), Spence (M.A.). 1974. The genetics of hypodontia. Journal of Dental Research 53, 781. Susanne (C.). 1977. Heritability of anthropological characters. Human Biology 49, 573-580. Suzuki (M.), Sakai (T.). 1954. On the “Protostylid” of the Japanese. Journal of the Anthropological Society Nippon 63, 81-84. Suzuki (M.), Sakai (T.). 1956. On the “Dryopithecus” pattern in recent Japanese. Journal of the Anthropological Society Nippon 44, 87-94. Suzuki (M.), Sakai (T.). 1973. Occlusal surface pattern of the lower molars and second deciduous molars among the living Polynesians. American Journal of Physical Anthropology 39, 305-316. Taffanel (O.), Taffanel (J.). 1957. La station préhistorique d'Embusco (commune de Mailhac, Aude). Cahiers Ligures de Préhistoire et d'Archéologie 6, 53-72. Tavares Da Silva (C.), Soares (J.). 1987. O povoado fortificado calcolitico do Monte da Tumba. Setubal Arqueologica 8, 29-79. Tavares Da Silva (C.), Soares (J.). 1998. Le Portugal. In: Guilaine (J.), ed. Atlas du Néolithique européen, 2B: l'Europe occidentale. Liège: Service de préhistoire de l’Université. Etudes et recherches archéologiques de l’Université de Liège. ERAUL 46, 997-1049. Tavares Da Silva (C.), Soares (J.), Cardoso (J.). 1995. Os povoados fortificados do Monte da Tumba e Leceia. Elementos para um estudo comparado. In: Kunst (M.), ed. Origens, Estruturas e Relaçoes das Culturas Calcoliticas da Peninsula Iberica. Actas das I Jornadas Arqueologicas (Torres Vedras, 3-5 abril 1987). Trabalhos de Arqueologia 7, 159-168. Tchérémissinoff (Y.). 2006. Les sépultures simples et plurielles du campaniforme et du Bronze ancien dans le Bassin rhodanien et ses zones d’influences Oxford: Archaeopress. BAR: International series S1531. Ten Cate (R.). 1998. Oral histology: development, structure, and function. St. Louis [etc.]: Mosby. Tenzcar (P.), Bader (R.S.). 1966. Maternal effect in dental traits of the house mouse. Science 152, 1398-1400. Thesleff (I.). 2003. Epithelial-mesenchymal signalling regulating tooth morphogenesis. Journal of Cell Science 116, 1647-1648. Thesleff (I.), Nieminen (P.). 1996. Tooth morphogenesis and cell differentiation. Current Opinion in Cell Biology 8, 844-850. Thoman (A.). 2000. Problèmes méthodologiques de quantification des individus d’une sépulture collective: Exemple du dolmen I des Cudières à Jouques (Bouches-du-Rhône). Université de la Méditerranée (AixMarseille II): Service d’anthropologie biologique (Diplôme d’études avancées non publié). Toth (K.). 2002. Kora bronzkori temetkezések Szeged-Kiskundorozsma hataraban. A Mora Ferenc Muzeum Evkönyve (Szeged). Studia archaeologica 8, 31-75. Toth (K.). 2003. A kora bronzkor kutatasanak helyzete Magyarorszagon. A Nyiregyhazi, Josa Andras Muzeum Evkönyve XLV évfolyam, 65-111. Townsend (G.C.), Brown (T.). 1980. Dental asymmetry in Australian Aboriginals. Human Biology 52, 661-673. Townsend (G.C.), Martin (N.G.). 1992. Fitting genetic Models to Carabelli Trait Data in South Australian twins. Journal of Dental Research 71, 403-409. 252 Bibliographie Townsend (G.C.), Yamada (H.), Smith (P.) 1986. The metaconule in Australian aboriginals: an accessory tubercle on maxillary molar teeth. Human Biology 58, 851-862. Townsend (G.C.), Yamada (H.), Smith (P.). 1990. Expression of entoconulid (sixth cusp) on mandibular molar teeth of an Australian aboriginal population. American Journal of Physical Anthropology 82, 267-274. Trancho (G.J.), Robledo (B.). 1999. Paleodieta: estudio del patrón alimenticio en El Cerro de la Cabeza (Avila). Junta de Castilla y León. Universidad Complutense Madrid. Trancho (G.J.), Robledo (B.), López-Bueis (I.), Fabián (F.J.). 1996. Reconstrucción del patrón alimenticio de dos poblaciones prehistóricas de la Meseta Norte. Complutum 7, 73–90. Tsai (P.L.), Hsu (J.W.), Lin (L.M.), Liu (K.M.). 1996. Logistic analysis of the effects of shovel trait on Carabelli's trait in a mongoloid population. American Journal of Physical Anthropology 100, 523-530. Tsuji (T.). 1958. Incidence and inheritance of the Carabelli’s cusp in a Japanese population. Japanese Journal of Human Genetics 3, 21-31. Tucker (A.S.), Matthews (K.L.), Sharpe (P.T.). 1998. Transformation of tooth type induced by inhibition of BMP signaling. Science 282, 1136-1138. Tucker (A.S.), Headon (D.J.), Courtney (J.-M.), Overbeek (P.), Sharpe (P.T.). 2004. The activation level of the TNF family receptor, Edar, determines cusp number and tooth number during tooth development. Developmental Biology 268, 185-194 Tummers (M.). 2004. To the root of the stem cell problem: the evolutionary importance of the epithelial stem cell niche during tooth development. University of Helsinki: Institute of biotechnology. (Dissertation non publiée). Turek (J.). 1995. Nálezy období zvoncovitých pohárů v povodí řeky Bíliny v severozápadních Čechach, The Finds of the Bell Beaker Period in the Bilina River region (North-West Bohemia). In: Blažek (J.), Meduna (P.), ed. Archeologické výzkumy v severozápadních Čechách 1983-1992. Most, 123-134. Turek (J.), Peška (J.). 2001. Bell Beakers Settlement in Bohemia and Moravia. In: Nicolis (F.), ed. Bell Beakers today: pottery, people, culture, symbols in prehistoric Europe: volume 2. International Colloquium (Riva del Garda, Trento, 11-16 May 1998). Trento: Servizio Beni Culturali, Provincia Autonoma di Trento, 411-428. Turek (J.), Dvořák (P.), Peška (J.). 2003. Archaeology of Beaker settlements in Bohemia and Moravia. An outline of the Current State of Knowledge. In: Czebreszuk (J.), Szmyt (M.), ed. The Northeast Frontier of Bell Beakers. Oxford: British Archaeological Reports. BAR: International series 1155, 183-208. Turner II (C.G.). 1967. Dental genetics and microevolution in prehistoric and living Koniag Eskimos. Journal of Dental Research 46, 911-917. Turner II (C.G.). 1969. Microevolutionary interpretation from the dentition. American Journal of Physical Anthropology 30, 421-426. Turner II (C.G.). 1970. New classifications of non-metrical dental variation: cusps 6 et 7. Paper presented at 39th annual meeting of the American Association of Physical Anthropologist. Washington D.C. Turner II (C.G.). 1981. Root number determination in maxillary first premolars for modern human populations. American Journal of Physical Anthropology 54, 59-62. Turner II (C.G.). 1985. Expression count: a method for calculating morphological dental trait frequencies by using adjustable weighting coefficients with standard ranked scales. American Journal of Physical Anthropology 68, 263-268. Turner II (C.G.). 1987. Late Pleistocene and Holocene population history of east Asia based on dental variation. American Journal of Physical Anthropology 73, 305-321. Turner II (C.G.). 1990. Major features of Sundadonty and Sinodonty, including suggestions about East Asian Microevolution, Population History, and Late Pleistocene relationships with Australian Aboriginals. American Journal of Physical Anthropology 82, 295-317. Turner II (C.G.), Hanihara (T.) 1977. Additional features of the Ainu dentition. V. Peopling of Pacific. American Journal of Physical Anthropology 46, 13-24. Turner II (C.G.), Scott (G.R.). 1977. Dentition of Easter Islanders. In: Dahlberg (A.A.), Graber (T.M.), ed. Orofacial Growth and Development. The Hague: Mouton Publishers, 229-249. Turner II (C.G.), Nichol (C.R.), Scott (G.R.). 1991. Scoring procedures for key morphological traits of the permanent dentition: the Arizona State University Dental Anthropology System. In: Kelley (M.A.), Larsen (C.S.), ed. Advances in dental anthropology. New York: Wiley Liss, 13-31. Tyrrell (A.J.). 2000. Skeletal non-metric traits and the assessment of intra- et interpopulation diversity: past problems and future potential. In: Cox (M.), May (S.), ed. Human osteology in archaeology and forensic science. London: Greenwich Medical Media LTD, 289-306. 253 Vaahtokari (A.), Åberg (T.), Jernvall (J.), Keränen (S.), Thesleff (I.). 1996. The enamel knot as a signaling center in the developing mouse tooth. Mechanisms of Development 54, 39-43. Van Valen (L.) 1962. A study of fluctuating asymmetry. Evolution 16, 125-142. Van Beek (G.C.). 1983. Dental morphology: an illustrated guide. Bristol: Wright. Vander Linden (M.). 2004. Elle tomba dans la bonne terre et elle donna du fruit qui monta (Marc 4, 8): la culture à céramique cordée comme substrat du phénomène campaniforme. Bulletin de la Société Préhistorique Française 101, 207-214. Vander Linden (M.). 2006. Le phénomène campaniforme dans l’Europe du 3ème millénaire avant notre ère. Synthèse et nouvelles perspectives. Oxford: Archaeopress. BAR: International series S1470. Vazquez Cuesta (A.). 2005. Alcohol y Campaniforme. Universidad de Navarra: Cuadernos de Arqueologia 13, 87114. Vazquez Cuesta (A.). 2006. Aproximacion al significado y uso de la ceramica campaniforme. Universidad de Alcala (Madrid): Departamento de historia y filosofia (Trabajo de investigacion de tercer ciclo non publié). Vaquer (J.). 1998. Le Mourral, Trèbes (Aude). A Fortified Languedocian Late Neolithic Site Reoccupied by Bell Beakers. In: Benz (M.), Van Willigen (S.), ed. Some new approaches to the Bell Beaker "Phenomenon": Lost Paradise...? Meeting of the Association Archéologie et Gobelets, 2 (Feldberg, Germany, April 18-20 1997). Oxford: British Archaeological Reports. BAR: International series 690, 15-21. Vencl (S.). 1994. K problému sídlišt’ kultury s keramikou šňůrovou. Archeologické Rozhledy 46, 3-24. Vladař (J.). 1989. Das Spätäneolithikum und die früheste Bronzezeit im nördlichen Karpatenbecken: ihre historische Bedeutung für die Entwicklung im eigentlichen Mitteleuropa. In: Buchvaldek (M.), Pleslová Štiková (E.), ed. Das Äneolithikum und die früheste Bronzezeit (C14 3000-2000 b.c.) in Mitteleuropa: kulturelle und chronologische Beziehungen. International Symposium, 14 (Prag-Liblice 20-24 oktober 1986). Prague: University Karlova, 223-225. Vokolek (V.). 1965. Pohřebiště zvoncovitých pohárů v Rosnicích-Das Gräberfeld der Glockenbecherkultur in Rosnice. Archeologické Rozhledy 17, 613-616. Vokolek (V.). 1981. Eneolitický hrob z Plotišť n. L. a pohřebiště zvoncovitých pohárů z Předměřic nad Labem-Ein archäologisches Grab aus Plotiště nad Labem und ein Gräberfeld der Glockenbecherkultur aus Předměřice n. L. Archeologické Rozhledy 33, 481-485. Von Burg (A.). 2002. Le Campaniforme sur le plateau de Bevaix. Archéologie Suisse 25, 48-57. Waddington (C.H.). 1957. The Strategy of the Genes. London: George Allen & Unwin. Wang (H.), Feng (G.Y.), Liu (W.Q.), Zhao (S.M.), Zhao (W.), Xue (H.), He (L.). 2000. Congenital absence of permanent teeth in a six-generation Chinese kindred. American Journal of Medical Genetics 90, 193-198. Wang (X.P.). 2004. Molecular mechanisms underlying tooth morphogenesis and cell differentiation. University of Helsinki: Institutes of biotechnology and of dentistry (Dissertation non publiée). Ward (J.H.). 1963. Hierarchical Grouping to optimize an objective function. Journal of the American Statistical Association 58, 236-244 Warmenbol (E.). 1996. Les débuts de l'Âge du Bronze en Belgique. In: Mordant (C.), Gaiffe (O.), ed. Cultures et sociétés du Bronze ancien en Europe. Colloque sur les Fondements culturels, techniques, économiques et sociaux des débuts de l'âge du Bronze. Congrès national des Sociétés savantes, 117, section de pré- et protohistoire (Clermond-Ferrand, 27-29 octobre 1992). Paris: Editions du Comité des travaux historiques et scientifiques (C.T.H.S.), 637-657. Weidenreich (F.). 1937. The dentition of Sinanthropus Pekinensis: a comparative odontography of the hominids. Peking: Paleontologica Sinica n.s. 1. Wheater (P.R.), Young (B.), Heath (J.W.). 2001. Histologie fonctionnelle. Bruxelles: De Boeck Université. White (T.D.), Folkens (P.A.). 2005. The Human Bone Manual. London: Elsevier Academic Press. Wilson (D.). 1851. The Archaeology and Prehistoric Annals of Scotland. London: Macmillan. Woelfel (J.B.), Scheid (R.C.). 1997. Dental anatomy: its relevance to dentistry. New-York: Williams et Wilkins. Wu (L.), Turner II (C.G.). 1993. Brief communication: variation in the frequency and form of the lower permanent molar middle trigonid crest. American Journal of Physical Anthropology 91, 245-248. Zápotocký (M.). 1960. Sídliste kultury zvoncovitych poháru u Kozel na Neratovicku-Die Siedlungsstätte der Glockenbecherkultur bei Kozly in Neratowitzer Gegend. Archeologické Rozhledy 51, 5-26. Zápotocký (M.). 1962. Nové mladoeneolitické hroby z Litoměřicka. Sborník Severočeského musea. Historia 3, 151162. Zápotocký (M.). 1986. Die Brandgräber von Vikletice-Ein Beitrag zum chronologischen Verhältnis von Stich-und Rhein-Bandkeramik. Archeologické Rozhledy 38, 623-649. 254 Liste des figures Chapitre 1 Figure 1. La céramique décorée campaniforme. Figure 2 . La céramique commune campaniforme. Figure 3. Le matériel associé au Campaniforme. Figure 4. Le matériel associé au Campaniforme. Figure 5. Le matériel associé au Campaniforme. Figure 6. Variabilité des structures funéraires à travers l’Europe. Figure 7. Variabilité des structures domestiques à travers l’Europe. Chapitre 2 Figure 8. Les régions étudiées dans ce travail. Figure 9. La Bohême, répartition des sites étudiés. Figure 10. La Hongrie, répartition des sites étudiés. Figure 11. Le sud de la France, répartition des sites étudiés. Figure 12. Le nord de l’Espagne, répartition des sites étudiés. Figure 13. La Suisse, répartition des sites étudiés. Chapitre 3 Figure 14. Anatomie de la dent. Figure 15. La dentition humaine. Figure 16. Les termes de positionnement. Figure 17. Présentation des différents systèmes employés pour la description des molaires supérieures. Figure 18. Présentation des différents systèmes employés pour la description des molaires inférieures. Figure 19. Présentation des différents systèmes de notation dentaire. Chapitre 4 Figure 20. Incisive latérale supérieure gauche présentant un développement de crêtes marginales sur les bords de sa surface linguale. Figure 21. Première molaire supérieure gauche montrant sur sa face mésiolinguale un tubercule accessoire plus connu sous le terme de tubercule de Carabelli. Figure 22. Quelques exemples de traits non métriques dentaires observés sur la dentition permanente. Figure 23. Le développement de la dent. Figure 24. Schéma représentant le modèle des champs morphogénétiques - Field theory - mis en place par P.M. Butler (1939). 255 Figure 25. Schéma reproduisant le modèle du Clone - Clone theory -, proposé par J.H. Osborn (1978), concernant le développement des dents postérieures (les molaires). Figure 26. Adaptation du modèle des variables à seuil de D.S. Falconer (1965) pour un trait non métrique hypothétique. Figure 27. Exemple de plaques de référence du système ASU (Arizona State University) reproduisant les différents stades que l’on peut observer dans l’expression des incisives en pelle. Figure 28. Exemple de plaques de référence du système ASU (Arizona State University) reproduisant les différents stades que l’on peut observer dans l’expression de certaines cuspides supplémentaires sur les molaires inférieures. Chapitre 5 Figure 29. Liste des traits non métriques dentaires observés sur la dentition permanente. Figure 30. Liste des traits non métriques dentaires observés sur la dentition déciduale. Figure 31. Le type des traits non métriques dentaires observés sur la dentition permanente. Figure 32. Le type des traits non métriques dentaires observés sur la dentition déciduale. Figure 33. Nombre total de dents enregistrées par observateur. Figure 34. Composition des différentes observations de la série de Villedubert. Figure 35. Décompte des déterminations communes discordantes selon leur origine et leur répartition. Figure 36. Evaluation des divergences absolues. Figure 37. Observed on one session only percentage. Variations des observations réalisées lors d’une session seulement. Figure 38. Variant scoring percentage. Evaluation des discordances de gradations d’un trait d’une session à l’autre. Figure 39. >1 grade variant scoring. Evaluation de l’attribution de plus d’un stade de différence. Figure 40. Net mean grade difference. Evaluation de l’erreur moyenne brute. Chapitre 6 Figure 41. Décompte des observations pour la Bohême. Figure 42. Décompte des observations pour la Hongrie. Figure 43. Décompte des observations pour le sud de la France. Figure 44. Décompte des observations pour le nord de l’Espagne. Figure 45. Décompte des observations pour la Suisse. Figure 46. Représentation synthétique traduisant l’état des données pour la Bohême. Figure 47. Représentation synthétique traduisant l’état des données pour la Hongrie. Figure 48. Représentation synthétique traduisant l’état des données pour le nord de la France. Figure 49. Représentation synthétique traduisant l’état des données pour le sud de l’Espagne. Figure 50. Représentation synthétique traduisant l’état des données pour la Suisse. Figure 51. Les différents types d’asymétrie liés à des interférences développementales définis par L. Van Valen (1962). Figure 52. Représentation synthétique touchant les résultats du degré de similarité de l’asymétrie des antimères pour la Bohême. 256 Figure 53. Représentation synthétique touchant les résultats du degré de similarité de l’asymétrie des antimères pour la Hongrie. Figure 54. Représentation synthétique touchant les résultats du degré de similarité de l’asymétrie des antimères pour le sud de la France. Figure 55. Représentation synthétique touchant les résultats du degré de similarité de l’asymétrie des antimères pour le nord de l’Espagne. Figure 56. Représentation synthétique touchant les résultats du degré de similarité de l’asymétrie des antimères pour la Suisse. Figure 57. La procédure suivie pour la fusion des données antimériques lors du comptage individuel individual count - mis en place par C.G. Turner et C.R. Scott (1977). Figure 58. Les dents clés fournissant selon C.R. Scott et C.G. Turner (1997) les meilleures informations dans les études entre populations. Figure 59. Représentation synthétique concernant les groupes de variables testés pour la mesure de dépendance intradistrict. Figure 60. Représentation synthétique concernant les groupes de variables testés pour la mesure de dépendance interdistrict. Figure 61. Représentation synthétique touchant les résultats du test de la dépendance du sexe dans l’expression des traits non métriques dentaires pour la Bohême Figure 62. Représentation synthétique touchant les résultats du test de la dépendance du sexe dans l’expression des traits non métriques dentaires pour la Suisse. Chapitre 7 Figure 63. Liste des formules utilisées pour chaque variable en vue de la transformation des incidences en fréquences uniques F.C. - frequence count. Figure 64. Schéma synthétisant la sélection des traits observés sur les dents clés qui seront intégrés dans la phase analytique. Figure 65. La Bohême, rappel de la localisation des sites. Figure 66. Représentation synthétique des dents-clés retenues pour les analyses de la Bohême (n=54) Figure 67. Profil pour chaque groupe testé dans les analyses, élaboré à partir des expression count des dents-clés retenues pour la Bohême (n=54). Figure 68. Résultats des analyses multivariées pour la Bohême. Figure 69. La Hongrie, rappel de la localisation des sites. Figure 70. Représentation synthétique des dents-clés retenues pour les analyses de la Hongrie (n=35). Figure 71. Profil pour chaque groupe testé dans les analyses, élaboré à partir des expression count des dents-clés retenues pour la Hongrie (n=35). Figure 72. Résultats des analyses multivariées pour la Hongrie. Figure 73. Représentation synthétique des dents-clés retenues pour les analyses des ensembles appartenant au domaine oriental (n=30). Figure 74. Résultats des analyses multivariées pour le domaine oriental. Figure 75. Le sud de la France, rappel de la localisation des sites. Figure 76. Représentation synthétique des dents-clés retenues pour les analyses du sud de la France (n=57). Figure 77. Profil pour chaque groupe testé dans les analyses, élaboré à partir des expression count des dents-clés retenues pour le sud de la France (n=57). Figure 78. Résultats des analyses multivariées pour le sud de la France. Figure 79. Le nord de l’Espagne, rappel de la localisation des sites. 257 Figure 80. Représentation synthétique des dents-clés retenues pour les analyses du nord de l’Espagne (n=17). Figure 81. Profil de chaque groupe testé dans les analyses, élaboré à partir des expression count des dents-clés retenues pour le nord de l’Espagne (n=17). Figure 82. Résultats des analyses multivariées pour le nord de l’Espagne. Figure 83. Représentation synthétique des dents-clés retenues pour les analyses des ensembles appartenant au domaine méridional (n=30). Figure 84. Résultats des analyses multivariées pour le domaine méridional. Figure 85. La Suisse, rappel de la localisation des sites. Figure 86. Représentation synthétique des dents-clés retenues pour les analyses de la Suisse (n=49). Figure 87. Profil pour chaque groupe testé dans les analyses, élaboré à partir des expression count des dents-clés retenues pour la Suisse (n=49). Figure 88. Résultats des analyses multivariées pour la Suisse. Figure 89. Représentation synthétique des dents-clés retenues pour les analyses Suisse et domaine oriental (n=30). Figure 90. Résultats des analyses multivariées Suisse et domaine oriental. Figure 91. Représentation synthétique des dents-clés retenues pour les analyses Suisse et domaine méridional (n=30). Figure 92. Résultats des analyses multivariées Suisse et domaine méridional. Figure 93. Représentation synthétique des dents-clés retenues pour les analyses entre les ensembles campaniformes (n=30). Figure 94. Profil pour chaque groupe testé dans les analyses, élaboré à partir des expression count des dents-clés retenues pour les ensembles campaniformes (n=30). Figure 95. Résultats des analyses multivariées pour les ensembles campaniformes. Figure 96. Représentation synthétique des dents-clés retenues pour les analyses sexes séparés (n=44). Figure 97. Profil de chaque groupe testé dans les analyses, élaboré à partir des expression count des dents-clés retenues pour l’étude sexes séparés (n=44). Figure 98. Résultats des analyses multivariées sexes séparés. Figure 99. Résultats des analyses multivariées sexes séparés avec intégration des groupes indéterminés. Chapitre 8 Figure 100. Dynamique des peuplements campaniformes. Synthèse de l’implication du fonds régional dans la mise en place du Campaniforme. Figure 101. Dynamique des peuplements campaniformes. Deux sphères populationnelles se distinguent. Figure 102. Dynamique des peuplements campaniformes de la phase 1 (environ 3000 - 2500 av. J.-C.) : diffusion au sein de la sphère méridionale. Figure 103. Dynamique des peuplements campaniformes de la phase 2 (environ 2500 - 2000 av. J.-C.) : diffusion au sein de la sphère orientale. 258 Liste des annexes Annexe 1. Catalogue des sites étudiés. Annexe 2. Catalogue des individus observés. Annexe 3. Catalogue des traits non métriques : dents permanentes. Annexe 4. Catalogue des traits non métriques : dents déciduales. Annexe 5. Fiches d’enregistrement manuelle des données : dents permanentes et déciduales. Annexe 6. Fiches d’enregistrement manuelle des données : dents isolées. Annexe 7. Exemple d’une fiche du formulaire de saisie. Annexe 8. Données fusionnées : la Bohême. Annexe 9. Données fusionnées : la Hongrie. Annexe 10. Données fusionnées : le sud de la France. Annexe 11. Données fusionnées : le nord de l’Espagne. Annexe 12. Données fusionnées : la Suisse. 259 Liste des tables Table 1. Evaluation de la concordance : la détermination dentaire - données brutes. Table 2. Evaluation de la concordance : les traits non métriques dentaires - données brutes. Table 3. Evaluation de la concordance : les traits non métriques dentaires - décomptes. Table 4. Evaluation de la concordance : les traits non métriques dentaires - NMGD. Table 5. Evaluation de la concordance : les traits non métriques dentaires - degré de similarité. Table 6. Statistiques descriptives : la Bohême. Table 7. Statistiques descriptives : la Hongrie. Table 8. Statistiques descriptives : le sud de la France. Table 9. Statistiques descriptives : le nord de l’Espagne. Table 10. Statistiques descriptives : la Suisse. Table 11. Corrélations antimères : la Bohême. Table 12. Corrélations antimères : la Hongrie. Table 13. Corrélations antimères : le sud de la France. Table 14. Corrélations antimères : le nord de l’Espagne. Table 15. Corrélations antimères : la Suisse. Table 16. Corrélations intradistricts : la Bohême. Table 17. Corrélations intradistricts : la Hongrie. Table 18. Corrélations intradistricts : le sud de la France. Table 19. Corrélations intradistricts : le nord de l’Espagne. Table 20. Corrélations intradistricts : la Suisse. Table 21. Corrélations interdistricts : la Bohême. Table 22. Corrélations interdistricts : la Hongrie. Table 23. Corrélations interdistricts : le sud de la France. Table 24. Corrélations interdistricts : le nord de l’Espagne. Table 25. Corrélations interdistricts : la Suisse. Table 26. Corrélations intertraits : tubercule de Carabelli (T24) - protostylide (T54). Table 27. Dimorphisme sexuel : la Bohême. Table 28. Dimorphisme sexuel : la Suisse. 261 Table 29. Incidence des traits par groupe - dentition permanente : la Bohême. Table 30. Incidence des traits par groupe - dentition déciduale : la Bohême. Table 31. Fréquences ajustées par la méthode de l’EC : la Bohême. Table 32. Variables et fréquences retenues pour l’analyse : la Bohême. Table 33. Incidence des traits - dentition permanente : la Hongrie. Table 34. Fréquences ajustées par la méthode de l’EC : la Hongrie. Table 35. Variables et fréquences retenues pour l’analyse : la Hongrie. Table 36. Variables et fréquences retenues pour l’analyse : le domaine oriental. Table 37. Incidence des traits - dentition permanente : le sud de la France. Table 38. Incidence des traits - dentition déciduale : le sud de la France. Table 39. Fréquences ajustées par la méthode de l’EC : le sud de la France. Table 40. Variables et fréquences retenues pour l’analyse : le sud de la France. Table 41. Incidence des traits - dentition permanente : le nord de l’Espagne. Table 42. Fréquences ajustées par la méthode de l’EC : le nord de l’Espagne. Table 43. Variables et fréquences retenues pour l’analyse : le nord de l’Espagne. Table 44. Variables et fréquences retenues pour l’analyse : le domaine méridional. Table 45. Incidence des traits - dentition permanente : la Suisse. Table 46. Incidence des traits - dentition déciduale : la Suisse. Table 47. Fréquences ajustées par la méthode de l’EC : la Suisse. Table 48. Variables et fréquences retenues pour l’analyse : la Suisse. Table 49. Variables et fréquences retenues pour l’analyse : Suisse-domaine oriental. Table 50. Variables et fréquences retenues pour l’analyse : Suisse-domaine méridional. Table 51. Variables et fréquences retenues pour l’analyse : les Campaniformes. Table 52. Incidence des traits - dentition permanente : CZ - sexe. Table 53. Fréquences ajustées par la méthode de l’EC : CZ - sexe. Table 54. Variables et fréquences retenues pour l’analyse : CZ - sexe. 262