Télécharger

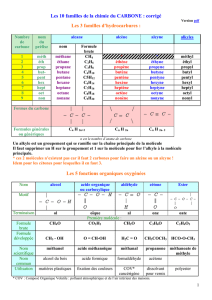
Ces différents modes d’alimentation ne sont connus que depuis
le milieu du XVIIesiècle. Il y a longtemps, on croyait que les végétaux
se nourrissaient exclusivement des matériaux bruts extraits du sol.
Vers la fin du XIXesiècle, les chercheurs savaient déjà que les végétaux
prélevaient de l’eau et du dioxyde de carbone pour les convertir, en
présence de lumière, en glucides et en oxygène. En 1883, le botaniste
T. Englemann voulut déterminer les longueurs d’onde de la lumière
solaire préférentiellement utilisées par les plantes.
Sachant que la photosynthèse libère de l’oxygène et que certaines
bactéries, comme la plupart des organismes, ont besoin de cette
molécule pour effectuer la respiration aérobie, Engelmann eut l’idée
d’éclairer différentes régions d’un organisme photosynthétique avec
tout le spectre de la lumière visible. Son hypothèse était que des
bactéries aérobies disposées le long d’une algue filamenteuse se dépla-
ceraient là où la production d’oxygène serait plus élevée, donc là où
l’activité photosynthétique serait la plus intense.
Il réalisa son expérience en déposant sur une lame microscopique
une goutte d’eau contenant des bactéries et y ajouta un filament d’algue
verte (Spirogyra). À l’aide d’un prisme de cristal, il sépara un faisceau de
lumière solaire et projeta la lumière décomposée sur la lame. Les bactéries
se multiplièrent principalement aux endroits où les algues vertes étaient
baignées de lumière violette et rouge, ce qui révéla l’efficacité de ces
longueurs d’onde pour stimuler la photosynthèse (voir la figure 7.1).
Non seulement ces observations illustrèrent-elles bien le fonction-
nement de la photosynthèse, mais aussi contribuèrent-elles à élucider
l’une des pierres angulaires de l’organisation du vivant: la photo-
synthèse est la principale voie par laquelle le carbone et l’énergie
pénètrent dans le réseau alimentaire des êtres vivants. Que les compo-
sés organiques soient fabriqués par un autotrophe ou absorbés par
un hétérotrophe, les cellules de cet organisme peuvent les utiliser
directement ou bien les emmagasiner. En effet, tous les organismes,
qu’ils soient autotrophes ou hétérotrophes, emmagasinent l’énergie
dans des composés organiques, pour ensuite la libérer au besoin par
des mécanismes tels que la respiration aérobie. La figure 7.2 présente
un aperçu des interactions chimiques qui existent entre la photo-
synthèse et la respiration aérobie, deux processus fondamentaux qui
font l’objet du présent chapitre et du suivant.
Ces processus peuvent sembler éloignés de nos préoccupations
quotidiennes. Cependant, il faut prendre conscience du fait que, sans
eux, les aliments de la plupart des organismes vivants, y compris les
humains, ne pourraient pas être produits et encore moins utilisés.
D’ailleurs, nous abordons ce point ultérieurement. L’étude de la
photosynthèse et de la respiration aérobie ouvre des perspectives sur
de nombreux enjeux d’importance tels que la nutrition, les régimes
alimentaires, l’agriculture, la croissance démographique, le génie géné-
tique, les impacts de la pollution sur les ressources agroalimentaires
et, par extension, la survie de l’humanité.
La lumière solaire et la survie
Lorsque nous ressentons la faim, nous éprouvons le besoin de manger.
Nous calmons alors notre faim en consommant des aliments de toutes
sortes: des fruits, des légumes, de la viande ou des pâtes, par exemple.
Cela dit, d’où viennent donc tous ces aliments ? Pour trouver la réponse,
nous devons regarder au-delà du réfrigérateur, du supermarché, du
restaurant ou même de la ferme. Les végétaux sont à l’origine de presque
tous les aliments que nous mangeons. Ils utilisent les sources d’énergie
et les matériaux bruts de leur milieu pour fabriquer le glucose et d’autres
composés organiques, constitués, rappelons-le, d’un squelette d’atomes
de carbone. On est dès lors en droit de se demander :
1. D’où viennent ces atomes de carbone?
2. D’où vient l’énergie qui alimente la synthèse des composés
carbonés ?
Les réponses à ces questions varient selon le mode nutritionnel de
l’organisme concerné.
Certains groupes d’organismes, incluant les végétaux, produisent eux-
mêmes les composés organiques qui les constituent à partir de matière
inorganique: ce sont des organismes autotrophes. Le carbone utilisé
est alors extrait du dioxyde de carbone (CO2), un composé gazeux
retrouvé dans l’atmosphère et dans l’eau. Les végétaux, certaines bac-
téries et de nombreux protistes sont photoautotrophes, c’est-à-dire
qu’ils extraient l’énergie du rayonnement solaire pour réaliser la photo-
synthèse. Au cours de ce processus, l’énergie solaire déclenche la
production d’ATP et de la coenzyme NADPH: l’ATP apporte de l’énergie
aux sites de la cellule photosynthétique où le glucose et les autres glu-
cides sont fabriqués, tandis que le NADPH les ravitaille en électrons et
en hydrogène, les éléments constitutifs des glucides en formation.
De nombreux autres organismes sont hétérotrophes: contrai-
rement aux végétaux, ils sont incapables de produire leur nourriture
eux-mêmes à partir d’énergie et de matériaux bruts tirés de leur envi-
ronnement. Ils doivent donc se nourrir d’organismes autotrophes,
d’autres hétérotrophes ou même de déchets organiques. C’est de cette
façon que la plupart des bactéries et de nombreux protistes survivent,
de même que les mycètes et les animaux.
7
COMMENT LES CELLULES SE PROCURENT L’ÉNERGIE
Figure 7.1 Les résultats de l’expérience de T. Engelmann établissent
une corrélation entre certaines longueurs d’onde de la lumière incidente
et l’efficacité de la photosynthèse chez Spirogyra, une algue verte filamen-
teuse. Un grand nombre de bactéries aérobies se sont déplacées vers
les régions où les algues avaient libéré le plus d’oxygène, ce dernier se
voulant un sous-produit de la photosynthèse.
Un prisme de cristal décompose le faisceau de
lumière en un spectre de couleurs projeté sur une
goutte d’eau déposée sur une lame de microscope.
Bactéries
(en blanc)
Algue filamenteuse
étalée sur toute la lame
Couleurs associées aux longueurs d’onde de la lumière (en nm)
22388_chap_07.qxd 3/13/06 3:51 PM Page 114

115
Figure 7.2 La relation entre la photosynthèse, le principal processus
biologique nécessitant de l’énergie, et la respiration aérobie, le principal
processus libérant de l’énergie.
PHOTOSYNTHÈSE
1. Le H2O est scindé par l’énergie
lumineuse. L’oxygène est libéré;
les électrons et l’hydrogène sont
utilisés par les chaînes de transport
qui libèrent l’énergie nécessaire à la
production d’ATP. Les coenzymes
captent les électrons et l’hydrogène.
2. L’ATP fournit l’énergie nécessaire
à la synthèse du glucose à partir
de l’hydrogène et des électrons fournis
par les coenzymes, ainsi que du car-
bone et de l’oxygène provenant
du dioxyde de carbone.
ATP disponible pour différents types de travail cellulaire
RESPIRATION AÉROBIE
1. Le glucose est décomposé
en dioxyde de carbone et en eau.
Les coenzymes captent ses électrons
et ses atomes d’hydrogène.
2. Les coenzymes cèdent des électrons
et de l’hydrogène à la chaîne de trans-
port d’électrons qui libère alors l’éner-
gie nécessaire à la production d’ATP.
Les végétaux
captent l’énergie
solaire qui alimente
la photosynthèse.
Concepts-clés
1. Les végétaux, certaines bactéries et beaucoup de protistes tirent
profit de l’énergie solaire, du dioxyde de carbone et de l’eau pour
fabriquer du glucose et d’autres glucides, qui sont tous des molécules
constituées d’un squelette carboné. Ce processus métabolique est
appelé photosynthèse.
2. La photosynthèse représente le principal processus par lequel le
carbone et l’énergie pénètrent dans la chaîne alimentaire.
3. Au cours de la première phase de la photosynthèse, la lumière
solaire est absorbée et convertie en énergie chimique emmagasinée
dans les molécules d’ATP. Des molécules d’eau sont scindées, leurs
électrons et leurs atomes d’hydrogène sont captés par la coenzyme
NADPqui devient alors du NADPH; l’oxygène, qui constitue le
sous-produit de ce processus, est libéré.
4. Lors de la seconde phase de la photosynthèse, l’ATP apporte
l’énergie aux sites de la biosynthèse. Les électrons et les atomes
d’hydrogène fournis par le NADPH se combinent au carbone et à
l’oxygène pour former du glucose. La source de carbone et d’oxygène
pour les réactions de synthèse se veut le dioxyde de carbone du milieu
environnant.
5. Chez les cellules végétales et chez les protistes photosynthé-
tiques, ces deux phases ont lieu au sein d’organites appelés chloro-
plastes.
6. L’équation suivante résume la photosynthèse :
12H2O 6CO26O2C6H12O66H2O
ÉNERGIE
LUMINEUSE
EAU DIOXYDE
DE CARBONE
OXYGÈNE GLUCOSE EAU
Eau,
dioxyde
de carbone
Oxygène
22388_chap_07.qxd 3/13/06 3:51 PM Page 115

Comment a-t-on découvert tout cela? En liant des radio-isotopes
aux atomes des réactifs illustrés dans l’équation globale (voir la sec-
tion 2.2), les chercheurs ont pu localiser dans les produits chacun
des atomes provenant des réactifs:
116 Chapitre 7 Comment les cellules se procurent l’énergie
Même par une journée chaude et ensoleillée, un humain
n’arrivera jamais à fabriquer lui-même la matière orga-
nique qui le compose. Alors comment les végétaux y
parviennent-ils? Ils doivent d’abord absorber l’énergie du
soleil pour ensuite effectuer une série d’opérations com-
plexes qui transforment l’énergie solaire en énergie chi-
mique qui sera, dans un premier temps, emmagasinée dans
les liaisons de l’ATP. Enfin, ils doivent se pro-
curer du carbone, de l’oxygène et de l’hydro-
gène, puis les combiner correctement
pour en faire des glucides grâce
à l’ATP. Il va sans dire que ces
tâches multiples ne peuvent être
exécutées au même site: cha-
cune d’entre elles relève d’une
structure particulière.
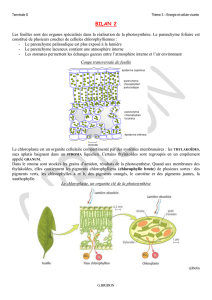
Le site des réactions
Chez les végétaux et les algues (des protistes), ces diffé-
rentes tâches sont exécutées dans le chloroplaste, un organite
spécialisé dans la photosynthèse (voir la section 4.7). La figure 7.3
illustre la structure typique d’un chloroplaste. Chacun est limité
par une double membrane qui entoure un liquide dense, le stroma.
Une autre membrane, souvent très repliée, baigne dans le stroma
et forme des empilements de sacs reliés par des canaux (voir la fi-
gure 7.3det e). Ces sacs, appelés thylakoïdes, s’empilent pour
former les grana. En fait, la membrane des thylakoïdes ne consti-
tue qu’un seul compartiment fonctionnel, peu importe le nombre
de replis qu’elle forme.
C’est dans les thylakoïdes que l’absorption de l’énergie solaire
et sa conversion en énergie chimique ont lieu. Grâce à des réactions
dépendantes de la lumière, appelées réactions photochimiques, les
végétaux obtiennent les électrons et les atomes d’hydrogène néces-
saires à la fabrication de leur nourriture. Lorsque la lumière solaire
atteint les chloroplastes, des molécules d’eau se scindent. L’oxygène
de l’eau diffuse dans l’atmosphère, tandis que ses électrons sont
transférés à des chaînes de transport d’électrons enchâssées dans la
membrane des thylakoïdes (voir la section 6.4). À mesure que les
électrons traversent la chaîne de transport, l’hydrogène est propulsé
à l’intérieur des thylakoïdes et, quand il en ressort, des molécules
d’ATP sont produites. Finalement, les électrons libres et l’hydrogène
sont captés par la coenzyme NADP, un accepteur d’électrons qui
devient alors du NADPH.
La synthèse du glucose, pour sa part, ne requiert pas de lumière;
elle se produit dans le stroma lors d’un cycle de réactions chimiques
utilisant l’énergie de l’ATP produit au cours des réactions photo-
chimiques. Cet ensemble de réactions indépendantes de la lumière,
appelé cycle de Calvin-Benson, incorpore les électrons et
l’hydrogène fournis par la coenzyme NADPH ainsi que le carbone
et l’oxygène du CO2.
Une équation simple peut servir à présenter globalement les
réactifs et les produits en jeu au cours de la photosynthèse:
12H2O 6CO26O2C6H12O66H2O
7.1
Figure 7.3 Un agrandissement des sites de la photosynthèse à l’intérieur
d’une feuille de plante typique.
La photosynthèse ne se termine pas avec
la synthèse du glucose
On présente le glucose comme étant le produit final de toutes ces
réactions. Cela résulte d’un choix arbitraire, qui sert à simplifier
l’explication de la séquence des réactions. En réalité, il y a peu de
RÉACTIFS
PRODUITS
UNE VUE D’ENSEMBLE DE LA PHOTOSYNTHÈSE
Face supérieure de la feuille Cellules photosynthétiques
bCoupe transversale d’un segment de feuille. Les régions
supérieure et inférieure contiennent un grand nombre de
cellules photosynthétiques.
cChloroplastes
dans une cellule
photosynthétique
(en vert)
Double
membrane
externe
Réseau membranaire
interne (thylakoïdes
reliés par des conduits) Coupe transversale d’un chloroplaste
aFeuille de laiteron
(Sonchus)
ÉNERGIE
LUMINEUSE
22388_chap_07.qxd 3/13/06 3:51 PM Page 116

P
NADPH
ADP
P
i
ATP
NADP
117
glucose dans les chloroplastes, car chaque nouvelle molécule de glu-
cose est rattachée à un groupement phosphate, ce qui la rend très réac-
tive. En général, elle se transforme rapidement en saccharose, en amidon
ou en cellulose, les principaux produits finals de la photosynthèse.
Le schéma ci-dessous se veut une version simplifiée de la figure 7.3.
Il sera utilisé dans d’autres sections de ce chapitre afin d’intégrer plus
facilement, au fur et à mesure, chaque détail du processus.
Avant d’aborder plus en détail certains aspects de la photosynthèse,
il est intéressant de noter que chaque chloroplaste est l’équivalent
d’une minuscule usine de production de sucres simples et d’amidon.
En effet, 2000 chloroplastes placés bout à bout ne mesurent que l’épais-
seur d’une pièce de 10 ¢. Il existe donc un nombre phénoménal de
chloroplastes dans un plant de maïs ou de riz. Cela révèle l’ampleur
des processus métaboliques requis pour jouer le rôle de producteur et
constituer la base des réseaux alimentaires (voir la section 48.1).
Les chloroplastes, les organites de la photosynthèse chez les végétaux
et les protistes photosynthétiques, sont spécialisés dans la produc-
tion de nourriture.
Au cours de la première phase de la photosynthèse, l’énergie
solaire alimente la production d’ATP et de NADPH. De l’oxygène
est alors libéré. Cette phase se déroule dans les chloroplastes,
plus précisément dans la membrane des thylakoïdes qui forme
un réseau parcourant le stroma.
La seconde phase de la photosynthèse se déroule dans le stroma.
En plus de l’énergie fournie par l’ATP, la synthèse du glucose
nécessite des atomes de carbone et d’oxygène (qui proviennent
du dioxyde de carbone), de même que des électrons et des atomes
d’hydrogène (fournis par le NADPH).
.
.
info.brookscole.com/starr10
H
2
O
CO
2
NADP
ATP
NADPH
e
e–
ADP + P
i
H
2
O
Lumière solaire Espace intrathylakoïdien
RÉACTIONS PHOTOCHIMIQUES
L’énergie solaire entraîne la scission des molécules
d’eau, une coenzyme capte l’hydrogène et les élec-
trons, puis l’ATP se forme.
Stroma
Canal
eReplis formés par la membrane des thylakoïdes
RÉACTIONS INDÉPENDANTES DE LA LUMIÈRE
Synthèse des glucides
Glucose
Saccharose, amidon, cellulose et autres
produits finals de la photosynthèse
fLes réactions photochimiques font intervenir des molécules qui captent la lumière,
des chaînes de transport d’électrons et d’autres composants. Tous ces éléments
sont enchâssés dans la membrane des thylakoïdes. Les réactions indépendantes
de la lumière, quant à elles, se déroulent dans le stroma.
Lumière
solaireAbsorption
d’eau
Absorption de
dioxyde de carbone
RÉACTIONS
PHOTO-
CHIMIQUES
RÉACTIONS
INDÉPEN-
DANTES DE
LA LUMIÈRE
Glucose
Libération d’eau
Libération d’oxygène
22388_chap_07.qxd 3/13/06 3:51 PM Page 117

118 Chapitre 7 Comment les cellules se procurent l’énergie
Les propriétés de la lumière
Le soleil est le point de départ d’un flux unidirectionnel d’énergie
qui se propage dans presque tous les écosystèmes (voir la figure 7.4).
Chaque mètre carré de surface terrestre reçoit en moyenne 7000 kcal,
et les photoautotrophes en interceptent environ 1%.
L’énergie du soleil voyage dans l’espace sous forme d’ondes électro-
magnétiques. La distance entre la crête de deux ondes successives cor-
respond à la longueur d’onde. La figure 7.5aillustre le spectre
électromagnétique, soit la gamme de toutes les longueurs d’onde
composant l’énergie rayonnante. Plus la longueur d’onde est courte,
plus l’énergie d’un rayonnement est élevée.
Figure 7.5
a) Le spectre électroma-
gnétique. La lumière visible
fait partie des nombreuses
formes de rayonnement
électromagnétique
du spectre. b) Un étudiant,
B. Mantoni, a créé un «arc-
en-ciel» en dirigeant
l’objectif de son appareil
photo vers un miroir.
La lumière du flash a été
réfléchie par le miroir vers
la lentille de son appareil,
qui a réfracté à différents
degrés, à la manière
d’un prisme, les rayons
des diverses longueurs
d’onde pour former
cet arc-en-ciel circulaire
imprimé sur la pellicule
photo.
a
7.2
400 450 500 550 600 650 700
Rayonnement de faible énergie
Rayonnement de haute énergie
b
Figure 7.4
Les photo-
autotrophes
constituent le
point d’entrée
dans la biosphère
du flux unidirec-
tionnel d’énergie
qui se propage
dans presque
tous les éco-
systèmes et les
réseaux d’êtres
vivants et dont
la source première
se veut le soleil.
Énergie
du soleil
Recyclage
des
nutriments
PHOTOAUTOTROPHES
(plantes, autres producteurs)
HÉTÉROTROPHES
(consommateurs, décomposeurs)
Libération d’énergie
(surtout sous forme de chaleur)
Rayonnement de
longueur d’onde courte
(les plus énergétiques)
Intervalle où se situent
la plupart des rayons
atteignant la surface terrestre
Intervalle où se situe la
chaleur libérée à la sur-
face de la Terre
Rayonnement de
longueur d’onde longue
(les moins énergétiques)
RAYONS
GAMMA RAYONS X RAYONNEMENT
ULTRAVIOLET PROCHE
INFRAROUGE RAYONNEMENT
INFRAROUGE MICRO-ONDES ONDES
RADIO
LUMIÈRE VISIBLE
Longueurs d’onde de la lumière (en nm)
LA LUMIÈRE SOLAIRE COMME SOURCE D’ÉNERGIE
Pour un grand nombre d’organismes, chaque longueur d’onde cor-
respond à une couleur. De plus, la lumière blanche que nous voyons
résulte de la combinaison de toutes les longueurs d’onde de la
lumière visible.
Collectivement, les photoautotrophes absorbent l’ensemble des
rayonnements de longueurs d’onde situées entre 380 et 750 nm. Par
ailleurs, les rayonnements de courtes longueurs d’onde, dont les
rayons ultraviolets (UV), sont suffisamment énergétiques pour briser
les liaisons chimiques des composés organiques; ils peuvent même
tuer les cellules. C’est pourquoi, il y a des centaines de millions
d’années, avant que la couche d’ozone se forme dans l’atmosphère,
le bombardement mortel des UV contraignait les organismes photo-
autotrophes à la vie sous-marine.
Nous savons que, lorsqu’un faisceau de lumière blanche traverse
un prisme de cristal (comme lors de l’expérience d’Engelmann), celui-
ci est décomposé en rayons de différentes longueurs d’onde déviés à
divers degrés, ce qui les fait diverger les uns des autres, formant ainsi
un spectre. Les arcs-en-ciel se forment de la même manière: les rayons
parallèles de la lumière solaire divergent les uns des autres au cours de
leur passage à travers les gouttelettes d’eau en suspension qui agissent
à la manière d’un prisme. En fait, un rayon lumineux se trouve d’autant
plus dévié par les gouttelettes qu’il possède une longueur d’onde longue.
C’est pourquoi les rayons de longueurs d’onde longues (de jaune à
rouge) divergent plus fortement de leur trajectoire d’origine que ceux
de longueurs d’onde courtes (de violette à bleu). Les couleurs se séparent
ainsi les unes des autres et forment un ensemble de cercles concen-
triques présentant dans l’ordre les couleurs du spectre, le violet étant
à l’intérieur et le rouge à l’extérieur (voir la figure 7.5b).
22388_chap_07.qxd 3/13/06 3:51 PM Page 118
 6
7
8
9
10
11
12
13
14
15
16
17
18
6
7
8
9
10
11
12
13
14
15
16
17
18
1
/
18
100%