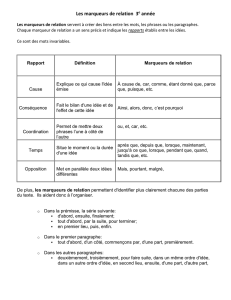
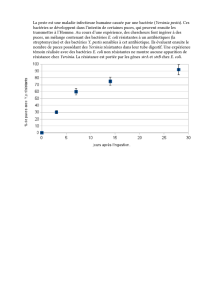
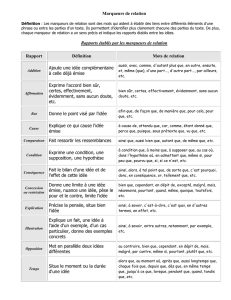
identification des sources de contamination - Archimer

Rapport d’activités de recherches
En vue de l’obtention du Diplôme
HABILITATION A DIRIGER DES RECHERCHES
Etude de la contamination microbiologique du milieu littoral :
identification des sources de contamination fécale et évaluation de
la persistance des bactéries entériques dans l’environnement.
Présenté par
Michèle Gourmelon
Autorisation de soutenance donnée par le conseil scientifique le 14/03/2013
Soutenance le 27 février 2014
Rapporteurs :
Mr. Anicet Blanch, Professeur, Université de Barcelone
Mr. Pierre Colin, Professeur émérite, ESIAB, Université Bretagne Occidentale, Plouzané
Mme. Fabienne Petit, Professeur, Université de Rouen
Examinateurs :
Mr. Frédéric Garabetian, Professeur, Université de Bordeaux I
Mr. Mohamed Jebbar, Professeur, Institut Européen de la Mer - Université de Bretagne
Occidentale, Plouzané
Mme. Anne-Marie Pourcher, Directeur de Recherche, Irstea, Rennes


Remerciements
Ce mémoire d’HDR résume le travail que j’ai réalisé au Laboratoire de Microbiologie, en
environnement littoral, à l’Ifremer, sur le centre de Brest au cours de plus d’une quinzaine d’années,
en collaboration avec des étudiants, les personnes du laboratoire et les partenaires scientifiques que
je tiens à remercier.
J’adresse tout d’abord tous mes remerciements à Anicet Blanch, Fabienne Petit et Pierre Colin
rapporteurs de ce mémoire ainsi qu’à Fréderic Garabetian, Anne-Marie Pourcher et Mohammed
Jebbar pour avoir accepté d’évaluer ce travail. La participation à ce jury sera peut-être le début de
futures collaborations.
Je tiens à remercier les étudiants qui ont contribué aux résultats présentés dans ce mémoire. Tout
d’abord, je remercie vivement Sophie Mieszkin, première étudiante en thèse que j’ai encadrée, qui a
fait un travail très conséquent au cours de sa thèse sur les traceurs de sources microbiennes et à qui
on doit de nombreux résultats sur les marqueurs Bacteroidales. Je tiens également à remercier
Aourell Mauffret qui a poursuivi ce travail en post-doctorat pour son dynamisme et les nombreuses
discussions. Merci également à Yustian Alfansiah, Charlotte Marin, Morgane Bougeard … Je tiens
aussi à remercier vivement Charlotte Balière qui est actuellement en thèse sur les E. coli producteurs
de Shiga-toxines au laboratoire.
J’adresse tous mes remerciements aux personnes du Laboratoire Santé, Environnement et
Microbiologie (SEM) de l’Ifremer, localisé à Brest et à Nantes. Je remercie tout d’abord Dominique
Hervio-Heath, responsable de l’équipe de Brest, pour son aide, ses conseils et discussions. Je
remercie également Marie-Paule Caprais, Jean-Claude Le Saux, Solen Lozach et Cécile Le Mennec
pour leur participation aux études réalisées tout au long de ces années, ainsi qu’Emmanuelle Quenot,
Joëlle Cozien et Noémie Flood qui sont arrivées plus récemment au laboratoire. Merci à tous pour
votre aide, votre bonne humeur et les bons gâteaux … Je tiens à remercier Monique Pommepuy,
responsable du laboratoire pendant de nombreuses années, qui m’a fait confiance pendant la thèse
et m’a ensuite permis de développer différentes thématiques au cours de ces années. Merci
également à Elisabeth Dupray et Annick Derrien pour leur participation à certaines études.
Je remercie également Soizick Le Guyader, responsable du laboratoire SEM pour ses conseils ainsi
que les autres collègues nantais : Pascal Garry, Françoise Hubert, Gaëlle Kaelin, Cédric Kergaravat,
Cécile Le Mennec, Estelle Le Quintrec, Jessica Maillot, Chantal Menanteau, Joanna Ollivier, Sylvain
Parnaudeau, Julien Schaeffer, Emilie Vallade, Antoine Véron … pour les discussions lors de nos
réunions et leur participation à des projets communs.
Je remercie également Tristan Renault, qui est responsable de l’Unité Santé Génétique et
Microbiologie des Mollusques dans laquelle s’insère notre laboratoire depuis janvier 2013. Ce
rattachement permettra des échanges très intéressants entre deux aspects différents de la
microbiologie en zone littorale. Je tiens également à remercier les nombreuses personnes des
Laboratoires Environnement Ressources de l’Ifremer qui nous ont apporté leur connaissance des
sites d’études et leur aide sur le terrain. Je remercie également les physiciens modélisateurs tels que
Pascal Lazure, Frank Dumas … avec qui nous avons collaboré dans le cadre d’études de modélisation
du devenir en mer des bactéries … Je remercie également les sédimentologues, chimistes ou

biologistes avec qui nous avons travaillé dans le cadre de projets pluridisciplinaires à l’Ifremer tels
que Philippe Bassoulet, Jacky L’Yavanck, Jean-François Guillaud …
J’adresse tout particulièrement mes remerciements à nos partenaires scientifiques qui ont permis la
réalisation d’une part importante du travail sur les traceurs de sources microbiennes au cours de ces
années : ceux de l’Irstea de Rennes : Anne-Marie Pourcher, Olivia Soleszki (post-doctorat), Romain
Marti (thèse), du CNRS Géosciences de Rennes : Emilie Jardé, Laurent Jeanneau, Loic Harault (post-
doctorat), de l’Université de Caen : Alain Rincé, de l’INRA de Narbonne : Nathalie Wéry, de
l’Université d’Angers : Alain Jadas-Hécart et Pierre-Yves Communal. Je tiens aussi à remercier nos
partenaires des laboratoires d’analyses des eaux : IDHESA, Gaël Durand, Hassiba Melikechi et
Emmanuelle Guillerm et Eurofins, Thierry Chesnot.
Je tiens à remercier également Gérard Corthier et Jean-Pierre Furet de l’Unité d’Ecologie et de
Physiologie du Système Digestif de l’INRA de Jouy en Josas pour les nombreux échanges au cours de
la thèse de Sophie Mieszkin.
Je remercie également nos partenaires impliqués dans la recherche des bactéries entériques
pathogènes tels que Christine Vernozy, Unité de Microbiologie Alimentaire et Prévisionnelle, Ecole
Nationale Vétérinaire de Lyon (actuellement à BioMerieux) et Delphine Thevenot et Estelle Loukiadis
d’AgroSupVet de Lyon et Martine Denis de l’ANSES de Ploufragan ...
Je tiens à remercier également nos collègues anglais avec qui nous collaborons depuis plusieurs
années et tout particulièrement Huw Taylor et James Ebdon de l’Université de Brighton et Jonathan
Porter de l’Environment Agency d’Exceter. Je remercie également Tom Edge de l’Environment
Canada, Tarja Pitkänen, The Finnish National Institute for Health and Welfare, Jorge Santo Domingo
de l’EPA de Cincinati pour les échanges et discussions.
J’adresse également tous mes remerciements aux personnes qui m’ont aidé pour la partie plus
administrative des projets et tout particulièrement les projets Interreg : Ramiro Gonzales, Helen Mc
Comby-Boudry, Marie-Christine Mazé, Joëlle Guillard, Véronique Gourmelon …
Je tiens également à remercier les financeurs de nos études : les programmes européens Interreg IIIb
et IVA, l’AFSSET, le Fonds Unique Interministériel, la Région Bretagne, l’Agence de l’eau Loire-
Bretagne, Brest métropole Océane, les Conseils Généraux du Finistère et de l’Ille et Vilaine, Cap
atlantique, l’Ifremer …
Je n’oublie pas non plus les amis : Véro, Aline, Emina, Laure, Fabrice, Georges, Olivier, Marie, Thierry,
Dany, Clément, Bradley … pour les bons moments de détente.
Je tiens à remercier mes parents et mes sœurs qui en assistant à la soutenance de cette HDR ont pu
ainsi avoir un aperçu de mes activités.
Enfin, un grand merci à Pascal, Fanny, Nicolas, Elise et Claire pour les bons moments en famille, leur
compréhension et soutien.

 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
1
/
113
100%