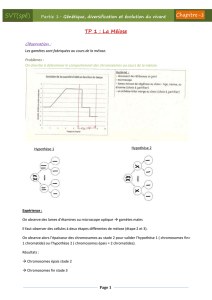
I - La reproduction sexuée des organismes

LSV1 – Semestre 2 – Biologie de la reproduction et du développement - 1
BIOLOGIE DE LA REPRODUCTION ET DU
DEVELOPPEMENT
LA REPRODUCTION SEXUEE DES
ORGANISMES SUPERIEURS
INTRODUCTION
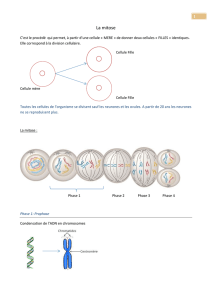
Il faut savoir que la mitose est un moyen de reproduction
asexuée des organismes unicellulaires. Elle amène à la
production de deux cellules filles, ayant un matériel
génétique identique l’une par rapport à l’autre et par
rapport à la cellule mère.
Il existe également des organismes pluricellulaires comme
l’hydre, qui se reproduisent via un mécanisme mitotique,
et dans ce cas là plus précisément sous la forme d’un
bourgeonnement cellulaire qui se détachera de
l’organisme parental. Leur patrimoine génétique sera
identique à moins que des mutations aient lieu lors de la
mitose.
Enfin, existe aussi un mécanisme de reproduction des
pluricellulaires par division de l’organisme en lui-même
puis régénération, comme les étoiles de mer.
Dans tous ces cas, l’organisme néoformé est toujours
identique à l’organisme parental. La reproduction asexuée
mène à la production de clones. Ce n’est évidemment pas
le cas lors de la reproduction sexuée.

LSV1 – Semestre 2 – Biologie de la reproduction et du développement - 2
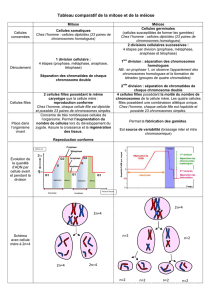
LA MEIOSE
La reproduction sexuée implique un mélange de deux génomes différents donnant naissance à des individus
qui contiennent chacun une combinaison unique de gènes. Ce mélange de génomes se fait par une union de
cellules haploïdes. Notre organisme, par exemple, est composé en grande partie de cellules diploïdes. Ainsi,
pour pouvoir se reproduire, il doit fabriquer des cellules haploïdes, qui donneront par union de nouvelles
cellules diploïdes. On a donc une réduction de ploïdie, qui a lieu lors de la méiose. « Meiosis » signifie en grec
réduction. L’haploïdie est relative à un seul jeu de chromosomes, et la diploïdie, deux jeux de chromosomes.
L’être humain possède 22 paires de chromosomes, des autosomes, plus deux chromosomes sexuels également
homologues.
Lors de la méiose, les cellules passent de deux jeux à un jeu de chromosomes. Les cellules formées sont
haploïdes et sont appelées gamètes. Ces deux gamètes vont se réunir, c’est la fécondation ou syngamie, pour
former de nouveau une cellule diploïde, le zygote. Le cycle de reproduction induit toujours une alternance
entre la production de cellules diploïdes et haploïdes selon le schéma suivant :
Méiose
cellules haploïdes
(1n)
Gamètes
(spermatozoïdes,
ovules)
Fécondation
Zygote (2n)
Cellules
germinales (2n)
Organisme
pluricellulaire
Organisme
pluricellulaire

LSV1 – Semestre 2 – Biologie de la reproduction et du développement - 3
CYCLE CHEZ L’HOMME ET LES ANIMAUX
Chez l’homme, les seules
cellules haploïdes sont
formées dans des tissus
spécialisés, les gonades,
où se trouvent des cellules
germinales, qui seront les
seules à subir une
réduction de ploïdie. Ces
gamètes ainsi formés vont
se réunir lors de la
fécondation pour former
le zygote, qui va se diviser
pour donner un organisme
pluricellulaire.
CYCLE CHEZ LES MYCETES ET LES ALGUES
Chez les mycètes et
certaines algues, le zygote
est la seule cellule diploïde.
Il est issu de la réunion de
deux cellules haploïdes, et
va tout de suite se diviser
via une méiose pour
donner naissance à deux
cellules haploïdes.

LSV1 – Semestre 2 – Biologie de la reproduction et du développement - 4
CYCLE CHEZ LES VEGETAUX ET CERTAINES ALGUES
Chez les végétaux et
certaines algues, le
zygote et les cellules
haploïdes sont capables,
chacun, de former un
organisme
pluricellulaire. Le zygote
va se diviser pour
donner un organisme
formé de cellules
diploïdes. Certaines de
ces cellules, par méiose,
vont donner naissance à
des spores, haploïdes,
qui peuvent se diviser
par mitose pour donner
naissance à un
gamétophyte. C’est ce
qu’on appelle une alternance de générations.
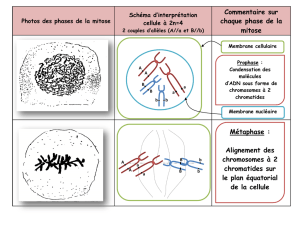
PROCESSUS MEIOTIQUE
Le processus de la méiose est le même chez les animaux et les végétaux. Ce processus méiotique implique une
réduction de ploïdie. La division cellulaire se situe après la phase G2 dans le cycle cellulaire.
1. En phase G1, les chromosomes sont constitués d’une seule chromatide. On a une cellule à 2n/2c.
2. Lors de la phase S, le matériel se duplique. Les chromosomes sont donc constitués de deux
chromatides sœurs. C’est donc une cellule diploïde à deux chromatides sœurs. la cellule est à 2n/4c.
3. En G2, la cellule est à 2n/4c.
4. La cellule rentre ensuite dans la division méiotique, qui va donner au final 4 cellules à 1n/1c.
La division méiotique est composé de deux divisions : la division réductionnelle ou méiose 1, qui réduit la
ploïdie, suivie de la division équationnelle, ou méiose 2. Lors de la méiose 1, les chromosomes homologues
vont être séparés en deux lots. On obtiendra deux cellules à 1n/2c. La méiose 2 ressemble beaucoup à une
division mitotique, et permet l’obtention de 4 cellules à 1n/1c. Elles constituent les gamètes.

LSV1 – Semestre 2 – Biologie de la reproduction et du développement - 5
MEIOSE 1
Elle est constituée d’une prophase 1, métaphase 1,
anaphase 1 et télophase 1.
LA PROPHASE 1
C’est une phase assez longue,
subdivisée en différents stades. Les
chromosomes homologues doivent au
final être séparés. Mais avant cela, ils
doivent se reconnaître. Cette
reconnaissance réciproque se fait lors
de la prophase 1. Se produit
également une recombinaison
génétique, un échange de segments
d’ADN entre des chromosomes non
sœurs mais homologues. Pour cela, les chromosomes
homologues doivent être au moins partiellement
identiques. La prophase peut occuper jusqu’à 80% du
temps total de la méiose. La répartition morphologique
des chromosomes permet la subdivision de la prophase
en 5 stades : leptotène, zygotène, pachytène,
diplotène, diacinèse.
STADE LEPTOTENE
Du grec « leptos », signifiant « fil ». Les chromosomes deviennent apparents
sous forme d’un « entremêlât » de fils, se font denses. On observe un
grossissement du noyau, les nucléoles sont visibles. On ne peut cependant
pas distinguer les chromosomes en eux-mêmes. Un assemblage d’axes
protéiques se forme le long des chromosomes, que l’on ne peut observer
en microscopie photonique.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
1
/
29
100%