L`influence favorable du foie embryonnaire sur Fhematopoiese in

/. Embryol. exp. Morph., Vol. 17,
2,
pp. 359-365, April
1967
359
With 2 plates
Printed
in
Great
Britain
L'influence favorable
du foie embryonnaire sur Fhematopoiese
in
vitro
dans
la
moelle osseuse de Fembryon de poulet
ParGERMANO SALVATORELLI1
Institut d'Embryologie experimental du College de France
et
du
C.N.R.S.,
Directeur: Professeur Etienne Wolff
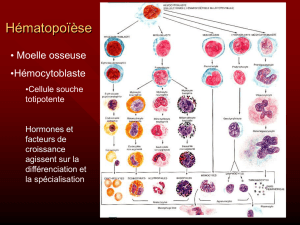
Pendant le developpement embryonnaire du poulet, on peut noter
la
succes-
sion de differents foyers d'erythropoiiese (Romanoff, 1960; Le Douarin, 1966):
(a)
les
ilots
de
Wolff dont l'activite
se
deroule entre
le ler et
2eme jour
d'incubation;
(b)
le
sac vitellin erythropoietiquement actif
du
4eme
au
19eme jour
d'in-
cubation;
(c) le foie et la rate dont la faible activite se deroule entre
le
7eme et le lOeme
jour d'incubation;
(d)
la
moelle osseuse dont l'activite commence au 12eme jour d'incubation et
qui deviendra chez l'adulte
le
seul organe ayant une activite erythropoietique.
On connait mal les facteurs necessaires
a
une erythropoiese normale, c'est-a-
dire
a
l'activation des cellules mesenchymateuses
et a
leur differentiation dans
les diverses lignees sanguines. De recentes recherches ont demontre
la
presence
de facteurs erythropoietiques dans le plasma. Mais, comme nous le verrons par
la suite, leur action ne peut pas expliquer entierement
le
processus de l'hemo-
poiiese.
La
methode
de
culture
in
vitro permet d'etudier dans des conditions
relativement simples les phenomenes de l'erythropoiese, mais il faut remarquer
que,
avec les methodes habituelles
de
culture, les differentes lignees sanguines
s'epuisent apres
24 a 48
heures
de
culture. Les cellules
qui
subsistent
et qui
peuvent etre cultivees plus longtemps
ne
sont
que des
elements
du
systeme
reticulo-endothelial.
La methode
de
culture
de
Wolff & Haffen (1952)
a
permis
au
contraire
de
garder pendant plusieurs jours de culture les differentes lignees sanguines, soit
des explants de moelle osseuse
de
poule (Pourreau-Schneider, 1962), soit des
explants de moelle osseuse humaine normale et leucemique (Pourreau-Schneider,
Bernard, Boiron
&
Wolff,
1963), soit d'aire vasculaire d'embryon
de
poulet
(Manelli, 1963; Raunich
&
Manelli, 1965; Raunich, Manelli, Mastrolia
&
Gardenghi, 1966), soit de foie embryonnaire de souris (Gallien-Lartigue, 1966).
1 Adresse de
Vauteur:
49 bis, avenue
de la
Belle Gabrielle, 94-Nogent S/Marne, France.
23 JEEM
17

360 G. SALVATORELLI
Nous avons done applique cette methode de culture a l'etude de l'hematopoiiese
medullaire pendant le developpement embryonnaire du poulet. Dans la presente
note,
nous exposons les resultats que nous avons obtenus en cultivant la moelle
osseuse d'embryons de poulet de 18 a 19 jours d'incubation, seule ou associee
a d'autres organes embryonnaires. A ce stade, les phenomenes de l'erythro-
poiiese sont en effet particulierement actifs (Sandreuter, 1951).
MATERIEL ET METHODES
La moelle osseuse de femur ou de tibia d'embryon de poulets de 18 a
19
jours
a ete explante"e selon la technique de Wolff
&
Haffen (1952) sur un milieu solide
recouvert de fragments de membrane vitelline d'oeuf de poule non incube
(Wolff,
1961). Le milieu solide se compose de: 10 gouttes de gelose en solution a
1 % dans le liquide de Gey; 4 gouttes de jus d'embryon de poulet de
8
jours et
demi d'incubation; 4 gouttes de serum de cheval.
En general, on n'a pas ajoute d'antibiotiques au milieu de culture. Nous
n'avons jamais eu d'infection dans nos cultures grace aux conditions de
rigoureuse asepsie dans lesquelles nous avons effectue nos experiences. Dans
une premiere serie experimentale, la moelle osseuse a ete cultivee seule; dans une
deuxieme serie, elle a ete associee au mesonephros d'embryons de poulet de 8
jours et demi; enfin dans une troisieme serie, au foie d'embryons de poulet de
7 a 11 jours.
Au total, on a cultive avec ces differentes techniques 1.200 explants environ,
dont une partie a ete fixee avec le melange de Helly ou de Bouin, et coloree au
Giemsa; l'autre partie a ete utilisee pour faire des frottis, colores ensuite par la
methode de May-Grunwald-Giemsa selon Pappenheim.
RESULTATS
(a) Moelle osseuse cultivee seule
La survie des explants ne depasse pas une semaine de culture. Pendant les
trois premiers jours de culture les explants gardent leur composition cellulaire
normale: la differentiation des cellules souches qui sont toujours presentes dans
nos explants, aussi bien que la maturation des elements, soit de la serie rouge,
soit de la serie blanche, continuent normalement.
A partir du 4eme jour de culture, on peut observer que la composition cellu-
laire des explants a subi une brusque variation: les erythrocytes montrent des
signes evidents de degenerescence; les erythroblastes deviennent de plus en plus
rares,
et, au 6eme jour de culture, ont presque disparu. Les cellules souches et
les elements de la serie granulocytaire, au contraire, meme apres 5 a 6 jours de
culture, sont tres nombreux et maintiennent leur capacite de se diviser par
mitose (Planche 2, figs. A, B).
Dans certains explants, on peut observer apres 4 a 5 jours de culture, la

J. Embryo/, exp. Morph., Vol. 17, Part 2 PLANCHE 1
Fig. A. Moelle osseuse associee au mesonephros apres 2 jours de
culture.
Les granulocytes ont
envahi les espaces intertubulaires. x 450.
Fig. B. Moelle osseuse associee au foie embryonnaire apres 14 jours de culture: presence
d'une
grande lacune vasculaire pleine d'elements sanguins. x 1125.
G.SALVATORELLIfacing p. 360

/. Embryol. exp. Morph., Vol. 17, Part 2
•#*
PLANCHE 2
B
. • «•
$
Fig. A. Moelle osseuse cultivee seule sur le milieu de culture apres 4 jours (frottis): une cellule
souche en mitose et quatre myelocytes. x 1400.
Fig. B. Moelle osseuse caltivee seule sur le milieu de culture apres
5
jours (frottis): un macro-
phage et un myelocyte en mitose. x 1400.
Fig. C. Moelle osseuse cultivee seule sur le milieu de culture apres 4 jours (frottis): trois
monocytes et un myelocyte. xl300.
Fig. D. Moelle osseuse associee avec du foie embryonnaire (frottis) apres
15
jours de culture:
un element non determine en mitose, deux erithrocytes et un granulocyte. x 1400.
Fig. E. Moelle osseuse cultivee associee avec du foie embryonnaire apres
15
jours (frottis): un
proerithroblaste et deux erithrocytes. x 1400.
Fig. F. Moelle osseuse cultivee associee avec du foie embryonnaire apres 15 jours (frottis):
deux erithroblastes, deux erithrocytes et un granulocyte. x 1400.
G.
SALVATORELLI
facing p. 361

Uhematopo'iese
in vitro 361
presence d'un nombre tres eleve de macrophages (Planche 2, fig. B), souvent
pleins d'elements sanguins phagocytes. Les elements monocytaires qui etaient
assez rares pendant les premiers jours de culture, deviennent, a partir du 3eme
jour, les elements cellulaires les plus frequents (Planche 2, fig. C).
(b) Culture de moelle osseuse associee au mesonephros
Dans ce cas aussi on ne peut maintenir l'hematopoiese normale pendant plus
de 3 a 4 jours de culture, et les resultats sont tout a fait comparables a ceux
qu'on obtient en explantant la moelle osseuse seule sur le milieu de culture.
Le mesonephros garde, apres 5 a 6 jours de culture, un aspect normal, tandis
que la moelle osseuse est caracterisee par un extreme appauvrissement dans sa
composition cellulaire: les elements de la serie rouge disparaissent bientot et
apres quelques jours de culture on ne trouve dans les explants que des cellules
souches et des elements de la serie granulocytaire et monocytaire. En ce qui
concerne le comportement du tissu myeloide vis-a-vis du tissu mesonephrique,
on peut noter que les deux tissus conservent en general leur individualite, mais
dans certains cas, quelques elements de la moelle osseuse, surtout des granu-
locytes, montrent un pouvoir d'invasion tres intense (Planche 1, fig. A). Us
colonisent les espaces intertubulaires et penetrent a l'interieur des tubes
renaux.
(c) Culture de moelle osseuse associee aufoie
En associant la moelle osseuse au foie embryonnaire, nous avons obtenu des
resultats tout a fait differents de ceux que nous avons obtenus avec les deux
premieres methodes de culture. Meme apres quinze jours de culture, la structure
et la composition cellulaire sont comparables a celles qu'on peut observer in
vivo.
A l'examen macroscopique les explants de moelle sont d'une couleur rouge
vif. Dans certains cas, on peut noter, a leur surface, un voile liquide de sang
nouvellement forme. L'examen microscopique des coupes nous montre l'exis-
tence de lacunes vasculaires remplies de cellules sanguines a differents stades de
leur maturation (Planche 1, fig. B). Le tissu hepatique maintient, pendant
toute la periode de culture, sa structure normale, a l'exception de sa partie
centrale ou Ton peut observer des phenomenes de necrose, surtout si l'explant
n'etait pas suffisamment mince. II faut aussi remarquer qu'on ne trouve jamais
dans le foie des centres d'hematopoiiese. La penetration des leucocytes de la
moelle osseuse dans le tissu hepatique est un phenomene tres rare a cause de la
structure compacte de son parenchyme. Les frottis de la moelle cultivee en
association avec le foie sont tres riches en elements soit de la serie rouge, soit de
la serie blanche (Planche 2, figs. D, E, F). Tous les stades de la differentiation
cellulaire depuis la cellule souche jusqu'a la cellule sanguine mure, sont presents
dans nos explants et dans des proportions comparables a ceux existant dans la
moelle osseuse normale (voir Tableau 1). Les erythrocytes qui avec les autres
23-2
 6
7
8
9
6
7
8
9
1
/
9
100%