Étude systématique des Tricorythidae afro

Étude systématique
des Tricorythidae afro-malgaches
(Insectes aquatiques, Ephéméroptère, Pan nota)
OLIARINONY Ranalison*
Département des Eaux et Forêts ESSA, Université d'Antananarivo, Madagascar * à
qui toute correspondance doit être adressée <ranalison.oliari[email protected]>
Résumé
Tous les stades de développement (œufs, larves, adultes) des
Tricorythidae (afrotropicaux et malgaches) ont été étudiés. Une nouvelle
espèce Madecassorythus raphaeli a été nouvellement découverte et
décrite. Les larves de Madecassorythus, Spinirythus, Ranorythus se
différencient de celles de Dicercomyzon et Machadorythus par la forme
générale du corps et des branchies. Le genre Tricorythus est le seul genre
ayant des yeux petits chez les mâles de Tricorythidae malgaches.
Introduction
Les Éphéméroptères font partie des insectes les plus anciennement connus de
l'homme. La première trace de l'existence des Éphéméroptères remonte à 9
siècles avant Jésus-Christ (SoldÂn, 1997). Aristote, environ 3 siècles avant J.-C,
fut le premier à signaler le caractère éphémère de la vie de ces insectes. Par
contre, la connaissance des éphéméroptères malgaches était extrêmement limitée
que jusqu'à une période récente (à partir de 1992) où le programme de recherche
sur la biodiversité et biotypologie des eaux continentales malgaches fut
développé.
Les Éphéméroptères jouent un rôle important à la chaîne trophique car ils
servent des nourritures des poissons.
L'ordre des Éphéméroptères comprend les sous ordres Schistonota et le sous
ordres Pannota. La famille Tricorythidae appartient à ce dernier sous ordre. Cette

0L1ARIN0NY 144
famille ne se rencontre qu'à Madagascar, en Malaisie (où il n'y a qu'une seule
espèce) et en Afrique orientale, dans la région éthiopienne.
En raison de la méconnaissance de la faune benthique afro-malgache, Me
Cafferty & De moor (1995a) ont souligné la priorité de l'étude systématique des
Éphéméroptères africains et malgaches. En plus, l'étude systématique est un outil
de base utilisé non seulement dans les diverses études en biologie et écologie,
mais surtout dans les études d'impact liées aux différents facteurs de perturbation
du milieu aquatique. Ainsi, les déchets chimiques provenant des industries ou bien
les engrais et pesticides utilisés en agriculture déversés à l'exutoire, quoique
encore à petite échelle à Madagascar, perturbent considérablement la santé de la
rivière et la vie de la faune qui s'y trouve. En outre, la déforestation de plus en
plus accélérée actuellement est la première cause de la dégradation du milieu
aquatique malgache. Dans ce cas, les Éphéméroptères sont classés parmi les bons
indicateurs de la qualité de l'eau à cause de leur sensibilité à la perturbation.
La systématique est aussi un outil de base pour la conservation car elle permet
aux écologistes de réfléchir aux mécanismes de l'évolution écologique de la
population (Funk & Piploy, 1990). Bref, toutes ces diverses raisons montrent
l'importance de l'approfondissement de l'étude systématique
Le but consiste à étudier la systématique de Tricorythidae malgaches à travers
une comparaison à la faune africaine. La révision de la famille est doublement
importante pour pouvoir confirmer la validité des taxa décrits précédemment
d'une part et pour apporter des compléments d'information d'autre part.
Méthodologie
La capture des larves a été réalisée au moyen d'un filet troubleau de vide de
maille de 250 um. Le prélèvement consiste à retourner les pierres pour récolter les
insectes à l'aide d'une pince souple ou un coup de brosse. Les larves au stade final

SYSIfcMAIIUUEDESTRICORYTTIDAE 145
reconnaissables à leurs ébauches alaires noires, sont mises en élevage dans un
cage de 10 cm de hauteur et de 6,5 cm de côté pour être observées afin d'obtenir
des compléments d'informations à leur détermination. La capture des imagos ou
des adultes a été réalisée à l'aide d'un filet à papillon d'ouverture de 40 cm de
diamètre et au moyen d'un piège lumineux composé de lumière blanche fournie
par une lampe à gaz et de lumière ultraviolette fournie par une petite lampe à UV.
Sur le terrain, les insectes sont conservés immédiatement dans un tube
contenant de Péthanol à 70%. L'étude (Suisse) des larves et des femelles a été
effectuée au Musée Cantonal de Zoologie de Lausanne. L'observation au
microscope à balayage a été réalisée au laboratoire de Microscopie électronique
au Centre des Hôpitaux et de l'Université de Lausanne (Suisse).
L'ensemble de matériel de cette étude est composé de la faune tricorythidienne
malgache. Les spécimens étudiés sont le fruit de 8 années de capture de 1991 à
1999. Quant à la faune africaine, elle est le résultat de 19 années de chasse de
1970 à 1989 en Afrique de l'Ouest, collectée lors de la réalisation du programme
de la lutte contre POnchocercose (collections données par Mr Jean-Marc
Elouard). Les spécimens sont déposés au Musée Cantonal de Zoologie à Lausanne
(Suisse).
La détermination des espèces au stade larvaire a été exécutée à l'aide d'un
microscope Olympus BH-2 après avoir monté les spécimens entre lame et lamelle
au baume de Canada. Chaque identification est basée sur l'observation de cinq à
six spécimens par prélèvement lorsque l'échantillonnage est suffisant.
En ce qui concerne les œufs, les femelles ont été disséquées. Quant au
montage, il s'agit de prélever les œufs conservés à l'éthanol à 70%, puis les placer
sur un turion métallique de 12 mm de diamètre. Le turion doit être mis dans une
boite hermétiquement fermée et déposé à l'étuve à 30°C pendant au moins 20 h
avant l'observation des œufs déshydratés. Pour observer la variabilité
morphologique des œufs au sein de l'individu ou au sein de la population, deux

OLIARINONY 146
turions par espèce ont été préparés, l'un porte les œufs provenant d'un seul
spécimen et l'autre les œufs de différents spécimens provenant des stations
différentes.
Avant l'observation au microscope à balayage (SEM) JSM-6300F, l'œuf a été
couvert d'une mince couche de platine de 90 Angstrôm. La métallisation suivie de
l'observation et la prise de photo dure régulièrement 4 heures par séance. En tout,
nous avons effectué 20 séances soit 80 heures d'observation et environ 180
photos.
Vu l'importance de l'effectif du genre Tricorythus (environ 870 larves de
Tricorythus africains et 1200 larves de Tricorythus malgaches), faute de temps,
nous nous sommes contentés d'étudier six espèces seulement dont trois malgaches
(T. ambinintsoae, T. sylvetsris et T. vulgaris) et trois africaines (T. discolor, T.
tinctus et T. lanceolatus).
Résultats
Toutes les espèces de Tricorythidae malgaches sont endémiques, en
provenance majoritairement de l'Est, des Hautes Terres, du Sud, de l'Ouest de
l'Ile et rarement du Nord. Parmi les 643 stations échantillonnées par l'équipe du
Laboratoire de Recherche sur les Systèmes Aquatiques et leur Environnements, la
présence de Tricorythidae est attestée dans 150 stations.
Tricorythus Eaton, 1868 •
Larves.
Aplaties dorso-ventralement. Yeux composés petits chez les deux sexes,
situés sur les extrémités postéro-latérales de la tête. L'ocelle médian est réduit,
nettement plus petit que les ocelles latéraux. Antennes : flagelle bien développé, 6
fois plus long que pédicelle. Labre : forme générale sub-rectangulaire, marge

SYSTEMATIQUE DES TRICORYTTIDAE 147
antérieure non échancrée, présence d'une ou plusieurs rangées des soies
courtes sur la partie antéro-latérale de la face ventrale. Mandibules : robustes,
canines externes formées de trois dents avec de courtes soies sur la partie distale,
canines internes à trois dents unies en cône ou une seule dent transformée en
forme de défense. Présence de soies fines entre les deux canines et sur la partie
médiane des canines internes. Prostheca allongée, pectinée dans la partie apicale,
avec quelques soies longues, plumeuses, bien dressées sur la mandibule gauche
mais rabattue sur la mandibule droite. Maxilles : galea-lacinia dépourvue de dents.
Présence de nombreuses soies longues sur la marge et la partie postéro-distale.
Palpe maxillaire de trois articles, le dernier court. Hypopharynx : marge antérieure
du lobe médian échancré, présence de soies fines sur la bordure apicale des lobes
latéraux. Labium : glosses et paraglosses soudées avec le pramentum, l'ensemble
formant une palette unique. Palpe labial de trois articles, l'apical très réduit, les
deux autres bien développés. Prothorax : chez les mâles, marge antérieure
convexe ou rectiligne, marge postérieure rectiligne tandis que chez les femelles,
les deux marges du prothorax sont sub-rectilignes. Pronotum deux ou trois fois
plus large que long. Pattes : pattes postérieures très développées, fémurs plus
élargis en comparaison des tibias grêles. Eperons coxaux sur PI, PII et PHI réduits
ou absents. Tibias longs surtout pour la patte antérieure. Tarses courts, griffe
tarsale avec quatre dents, deux basales et deux apicales. Abdomen : segments
abdominaux VII à IX plus longs que les autres. Branchies : 5 paires présentes sur
les segments II à VI, de taille décroissante, formées de deux lamelles ; la
supérieure ovoïde et l'inférieure constituée de deux séries de diverticules. Lamelle
inférieure bien développée, plus longue que la supérieure. Ebauches génitales :
toujours à l'intérieur. Filaments caudaux : paracerque plus long que les cerques.
Chaque segment est entouré d'un verticille d'épines allongées. Il faut noter que les
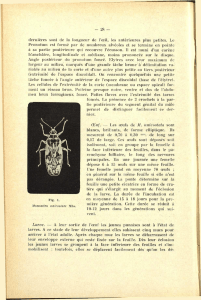
schémas d'une larve ainsi que les pièces buccales montrés aux Fig. 1 et Fig. 2 sont
extraits des travaux ultérieurs.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
1
/
30
100%