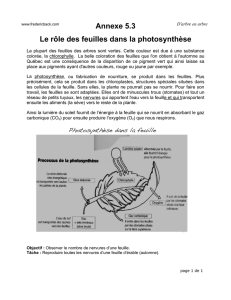
La Nutrition Carbonnée

La Nutrition Carbonée.
1. Introduction
La photosynthèse est un mécanisme fondamental qui permet la conversion de l’énergie
lumineuse en énergie chimique.
Il existe un angle écologique car c’est un processus à l’origine de la fabrication de la
biomasse terrestre. On a conversion d’une forme de carbone la plus oxydée en forme plus
réduite : CO
2
sucres, protéines, lipides.
S’il n’y a pas de photosynthèse, il n’y a pas de biomasse et, la photosynthèse produit de
l’O
2
qui représente 20% des composants de l’atmosphère.
Les végétaux sont autotrophes. Ils se nourrissent à partir de formes non réduites de
carbones, eau, oxygène… Leur capacité hétérotrophique est moins importante, ils utilisent des
carbones organiques tel que l’urée mais, pendant la germination, les plantes sont
hétérotrophes.
Les plantes peuvent assimiler le CO
2
lors de la transformation du pyruvate en
oxaloacétate grâce à la pyruvate carboxylase, c’est la β-carboxylation. On a une simple
fixation du CO
2
sans lumière.

2. Historique
• Rejet d’oxygène
Priestley a montré qu’une plante peut vivre sous une cloche en verre sans
renouvellement d’air contrairement à un animal.
Les plantes produisent de l’O
2
(1771-1773)
• Lumière indispensable (1779)
• CO
2
absorbé en même temps que l’émission d’O
2
(Senestien 1782-1783)
• La lumière agit comme source d’énergie (Mayer 1845)
• Chloroplastes importants (Sachs 1864)
CO
2
+ H
2
O + Energie Lumineuse Glucides + O
2
• Dégagement d’O
2
indépendant de la fixation du carbone (Hill 1937)
Réaction de Hill : Chloroplastes isolés + agents oxydants (accepteurs d’électrons)
production d’O
2
L’intégrité des chloroplastes n’est pas nécessaire. Seuls les thylakoïdes sont
nécessaires.

La réaction globale est une réaction d’oxydo réduction. D’où viennent les électrons ?
Les atomes d’oxygènes ?
Hill suggère que
H
2
O libère des électrons et de l’
O
2
en se dissociant.
• Ruben et Komen (1940) démontrent que l’O
2
provient de l’H
2
O de même que les
électrons libérés.
• Van Niel (1941) démontre avec des bactéries photosynthétiques sulfureuses que le
donneur d’électron peut ne pas être de
H
2
O :
CO
2
+ 2H
2
S HCHO + H
2
O + 2S
On a une différence entre les photosynthèses oxygéniques (production d’O
2
) et les
photosynthèses anoxygéniques (pas de production d’O
2
).
Principe thermodynamique : tout système évolue spontanément d’un état moins probable à
un état plus probable.
C CO
2
: forme la plus oxydée, la plus stable.
3. Structure et ultrastructure des chloroplastes
Ce sont des organes allongés. Il en existe plusieurs dizaines par cellules chez les
végétaux supérieurs. Leur nombre et leur taille varient en fonction de l’âge de la cellule. On les
trouve dans les parties aériennes des plantes : tiges et feuilles.
Les stomates possèdent des chloroplastes.
Energie
Lumineuse
Réaction
Lumineuse
(thylakoïdes)
H
2
O CO
2
Fixation
Carbone
(Stroma)
NADPH
ATP
H
2
O CO
2

Les chloroplastes sont issus de la différenciation des proplastes. Ils sont d’origine
maternelle (hérédité cytoplasmique). Toute la machinerie de transcription de l’ADN existe chez
les plastes qui contiennent de l’amidon, des globules lipidiques.
Les chloroplastes sont le lieu de synthèse des acides gras, des caroténoïdes.
Ils sont délimités par une enveloppe constituée par une double membrane. Cette
enveloppe limite l’espace intérieur, le stroma où se trouvent les enzymes nécessaires aux
diverses réactions.
Dans le stroma on trouve les thylakoïdes qui sont des empilements de saccules aplaties
appelées grana où on trouve les réactions de Hill. La lumière du thylakoïdes est appelée
espace inter membranaire.

Il existe des thylakoïdes courts qui forment les grana et des thylakoïdes longs qui relient
les grana entre eux. Ils sont parallèles à l’axe du chloroplaste. Ces thylakoïdes contiennent la
chaîne de transport d’électrons, les pigments chlorophylliens et le complexe ATP synthétase.
Plus il y a d’empilements et mieux la récupération d’énergie lumineuse se fait. Une
plante à l’ombre à plus de grana qu’une plante en lumière forte.
4. Molécules impliquées dans la photosynthèse
a. Pigments
On distingue deux pigments :
- Les caroténoïdes
-
Les chlorophylliens associés à des protéines. On parle de chloroprotéines.
-
La chlorophylle est constituée d’un noyau porphyrine : 4 pyroles en cercle et un atome de
magnésium chélaté au centre. On a associé à ce noyau un alcool à large chaîne appelé
phytol.
Chlorophylle a : pigment bleu vert présent chez tous les végétaux
Chlorophylle b : pigment vert jaune chez les plantes et les algues vertes
Chlorophylle c/d : chez les algues brunes et rouges.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
1
/
44
100%