Echanges hémato tissulaires

27/02/06 Physiologie de la respiration Abel et Adélaïde
V/ ECHANGES GAZEUX HEMATO-TISSULAIRES
A/ Définition et mécanismes des échanges
Se sont les échanges de gaz entre le sang et la circulation capillaire tissulaire et les cellules sans
préjuger du sens de ces échanges : oxygène du sang vers les cellules
CO2 des cellules vers le sang capillaire.
C’est le même mécanisme qu’au niveau alvéolo-capillaire. C’est un phénomène passif,
physique, de diffusion en fonction d’un gradient de pression partielle : un gaz va de l’endroit où
sa pression est la plus élevée vers l’endroit où sa pression est la plus basse(l’O2 quitte le sang et
le CO2 gagne le sang).
B/ Facteurs conditionnant les échanges
1-PRESSION TISSULAIRE DES GAZ
Dans les poumons, on a : V=Dl*[Pa-Pc]
Dans les tissus, on a : V=[Pc-Pt]*Dl
Dans les tissus, les pressions moyennes sont faibles voir très faibles mais au contact des
mitochondries, on a 3 à 5 mmHg de pression.
C’est pourquoi la pression Pc est toujours supérieure à la pression Pt donc il y a toujours des
échanges(il faudrait une pression quasi nulle dans les capillaires pour avoir un arrêt des
échanges hémato-tissulaires).
L’O2 est transporté par les globules rouges. Si la pression est toujours suffisante pour les
échanges, l’oxygénation dépend du contenu artériel en O2 : quand on a une anémie, on a une
diminution des globules rouges et donc moins d’O2.
2-VASCULARISATION DES TISSUS
La membrane représente la surface des capillaires et définit la distance que doit parcourir
l’oxygène.
a) Distance de diffusion
On a deux façons de la concidérer :
- on prend une cellule et on regarde à quelle distance elle se trouve d’un capillaire actif.
Sur le premier schéma de la huitième feuille de schéma :
la cellule 2 est très proche des capillaires
la cellule 1 en est éloignée donc elle aura du mal à être oxygénée.
La cellule 3 est au contact d’un capillaire fermé donc elle est dans la même situation que la
cellule 1.

- la distance entre deux capillaires est adéquate pour fournir de l’oxygène à l’ensemble du
tissu entre ces deux capillaires.
Quand la consommation en oxygène augmente, on peut atteindre un stade critique où la
distance est juste suffisante pour oxygéner les cellules les plus éloignées.
Quand la distance est trop importante, les premières cellules les plus proches des capillaires se
servent abondamment et les cellules les plus éloignées ne reçoivent plus d’oxygène donc elles
fonctionnent en anaérobie en faisant de l’acide lactique, cela n’a lieu que quand le tissu est très
actif et que la vascularisation ne suit pas.
b)Surface de diffusion
Un capillaire est un cylindre, les échanges se font sur toute la surface des capillaires qui
représente : au repos->150m2
en exercice physique->1000m2
la totalité des capillaires de l’organisme->6000m2.
Quand on fait un exercice physique, on recrute des capillaires et on augmente d’autant la
surface pour les échanges entre les cellules et le sang.
La modification des métabolites locaux entraîne l’ouverture des capillaires, leur recrutement.
La diminution de la Po2,
l’augmentation de la Pco2,
l’augmentation de la température,
la diminution du pH,
l’augmentation de l’ADP,
l’augmentation de l’acide lactique et
l’augmentation du K+ entraînent une augmentation de la surface de diffusion pour
favoriser les échanges.
c)Conséquences physiopathologiques
La Pao2 conditionne les échanges et le contenu artériel en oxygène conditionne le volume des
échanges. Ce qui joue, c’est la vascularisation des tissus.
Si la Pao2 et la Caco2 sont normales ainsi que la vascularisation alors tout est normal sauf si
on utilise certains poisons comme le cyanure qui empêche les échanges.
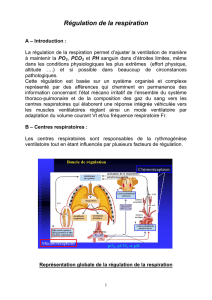
VI/ COMANDE ET ADAPTATION DE LA VENTILATION PULMONAITRE
A/ Commande de la ventilation pulmonaire
1- NEUROGENESE VENTILATOIRE
La ventilation peut être modifiée durant certaines situations : la parole, la déglutition,
l’exercice musculaire, …
Il y a des mécanismes qui règlent la fréquence, le rythme et l’amplitude de la ventilation par le
système nerveux. Ce système est basé sur des centres respiratoires dans le système nerveux

central qui envoient des efférences vers des effecteurs qui sont les muscles respiratoires pour
adapter la ventilation aux besoins. Les infos sont reçues par des récepteurs puis empreintent les
voies afférentes.
2-CENTRES RESPIRATOIRES
a) Localisation
La connaissance de cette localisation provient des expériences menées chez l’animal et chez
l’homme anesthésié, or on ne sait pas si l’anesthésie change quelque chose.
On a des centres au niveau bulbaires = centres régénérateurs de la ventilation, ils sont pairs et
symétriques : groupe respiratoire dorsal et
groupe respiratoire ventral.
au niveau pontique = centre pneumotaxique qui a un rôle modulateur.
° Le groupe respiratoire dorsal reçoit toutes les afférences qui apportent les informations
périphériques pour réguler les besoins : infos apportées par les nerfs IX et X.
Ce groupe envoie des informations au centre pneumotaxique et au groupe respiratoire ventral.
Ce groupe est le seul à recevoir des informations mais il participe également à la ventilation car
il contient des neurones pour des muscles, uniquement ceux inspiratoires et plus
particulièrement au seul diaphragme. Les informations partent en contro-latéral.
On a deux groupes de neurones inspiratoires :
- ce qui déchargent en tout début et même avant l’inspiration : se sont les
«early I »,
- ce qui décharge pendant toute l’inspiration : se sont les générateurs de rampe,
ils augmentent leur décharge durant toute l’inspiration.
Le groupe respiratoire dorsal contient ses deux types de neurones.
° Le groupe respiratoire ventral contient deux noyaux :
- A = ambigu,
- RA = rétro-ambigu.
Le noyau A a sous sa dépendance toutes les voies aériennes et les informations du groupe
respiratoire dorsal.
Le noyau RA envoie des neurones respiratoires en contro-latéral pour le diaphragme et les
intercostaux externes = muscles inspiratoires,
Ce noyau contient des neurones expiratoires qui sont comme les neurones inspiratoires early I =
early E et
d’autres qui déchargent durant toute l’expiration mais qui ne sont pas
générateur de rampe.
Le noyau RA est en connexion le noyau RA de l’autre coté pour favoriser la synchronisation
de la respiration.
° Le noyau pneumotaxique reçoit des informations du groupe respiratoire dorsal.
b)Mode de fonctionnement
Il y a plusieurs hypothèse :
- il existe une oscillation bi-stable qui est une inhibition réciproque et alternative.
En début d’inspiration, les early inspiratoires inhibent les neurones expiratoires et l’inspiration
se déroule donc avec les neurones inspiratoires, les autres sont inhibés. Au bout d’un certain

temps, les neurones expiratoires s’accommodent et s’échappent, en premier, se sont les early
expiratoires s’échappent et inhibent les neurones inspiratoires, puis ils vont s’accommoder et
s’échapper pour de nouveau inhiber les neurones expiratoires, …
C’est au niveau des centres bulbaires que réside l’automatisme de la ventilation.
Le centre pneumotaxique est un modulateur, il modifie le seuil d’excitabilité du système
d’interruption de l’inspiration. Il s’assure qu’après l’inspiration, il y a l’expiration durant un
temps suffisamment long.
B/Adaptation de la ventilation pulmonaire
1-MISE EN JEU DES MECANISMES ADAPTATIFS
a) Réflexe
C’est la mise en jeu d’un effecteur à la suite de la stimulation d’un récepteur sensitif ou
sensoriel.
° - Chémorécepteurs artériels :
Se sont les principaux éléments de la réponse réflexe.
Ils sont au niveau de la crosse de l’aorte et des carotides. Ces chémorécepteurs sont en
dérivation de l’artère, dans des corpuscules qui sont des glomi, ces glomi contiennent des
récepteurs qui dérivent du système Amine Precursor Uptake Decarboxylating (apud). Les
cellules de ce système sont des chémorécepteurs en dérivation de l’artère pour réguler la
ventilation.
Les chémorécepteurs sont sensibles :
- aux variations de Po2 : une diminution de la Po2 entraîne une augmentation
de la ventilation
- aux variations de la Pco2 : une augmentation de la Pco2 entraîne une
augmentation de la ventilation
- aux variations du pH : une diminution du pH entraîne une augmentation de la
ventilation.
Ces informations sur la Po2, la Pco2 et le pH sont des influx excitateurs pour la ventilation,
elles cheminent sur les fibres nerveuses du glomus carotidien puis dans le nerf de Herring puis
dans le nerf glosso-pharyngien,
fibres nerveuses du corpuscule aortique puis dans le nerf de Cyon puis
dans de nerf vague.
Les afférences sont des fibres nerveuses qui cheminent dans les même voies que celles des
barorécepteurs.
°- Récepteurs broncho-parenchymateux :
Ces récepteurs renseignent les centres respiratoires sur ce qui se passent dans les poumons. Les
afférences passent par le nerf vague. Les récepteurs des voies aériennes supérieures sont dans
l’épithélium : soit sous les cellules
soit entre les cellules.
Ces récepteurs sont sensibles aux particules inhalées,
aux gaz irritants,
aux corps étrangers.

Quand ces récepteurs sont stimulés, la réponse est de type : secousse de toux, constriction
laryngé et trachéale.
On utilise un fibroscope pour voir ce qui se passe dans les bronches mais pour cela, il faut
anesthésier les endroits où l’on passe pour éliminer ce réflexe.
On a, en plus, des récepteurs à l’irritation des bronches plus petites qui se trouvent sous ou
entre les cellules de l’épithélium. Ils sont sensibles à l’air froid.
Quand ils sont stimulés, on a une hyperventilation et une bronchoconstriction mais pas de
secousse de toux.
Il existe le réflexe de toux paradoxal de Head :
c’est un renforcement de l’effort inspiratoire à la suite d’inspirations profondes = soupir.
On trouve, également, des récepteurs à l’étirement.
Quand les bronches se dilatent et que les alvéoles se gonflent, la réponse de ces récepteurs qui
se trouvent dans les fibres musculaires lisses, est une diminution de l’inspiration du fait de la
dilatation de la bronche lors de l’inspiration -> c’est le réflexe de Herring-Bereuer qui ne
marche pas au repos.
Ce réflexe existe chez le nouveau-né
durant l’exercice physique quand Vt est supérieur à 1L et
réapparaît en pathologie durant l’emphysème pulmonaire quand la distension
est trop importante.
Il existe des récepteurs juxta-capillaires qui se trouvent dans le parenchyme entre les alvéoles
et le capillaire. La stimulation est provoquée par la distension du capillaire et par la présence
anormale de liquide dans le tissu interstitiel, elle entraîne une respiration rapide et superficielle
jusqu’à l’apnée quand les récepteurs sont très stimulés. Ces récepteurs seraient à l’origine des
signes responsables de cardiopathies.
°- Récepteurs musculaires :
Ils se trouvent dans les muscles striés squelettiques :
- respiratoires où il y en a de deux sortes -> les fuseaux neuro-musculaires qui
renseignent sur la longueur des muscles et
-> les organes tendineux de Golgi
pour informer sur la force développée par les muscles.
Ils ont un rôle majeur dans la stabilité ventilatoire : ils sont à l’origine de la capacité à
maintenir une ventilation adaptée malgré les interférences mécaniques de la cage thoracique.
- muscles de l’appareil locomoteur au niveau des tendons et des articulations.
Ils sont à l’origine de l’accrochage et du décrochage ventilatoire durant l’exercice physique.
Durant l’exercice physique, la ventilation augment pour augmenter l’oxygénation musculaire :
on a une augmentation brutale qui représente l’accrochage et quand on arrête l’exercice, on a
une diminution brutale du débit ventilatoire qui est le décrochage. Dès que l’articulation bouge,
les récepteurs envoient des influx qui sont des réflexes anticipateurs.
°- Autres récepteurs :
Il existe : - des barrorécepteurs : quand la PSA augmente, la ventilation diminue par des influx
inhibiteurs. C’est une réponse mineure dans cette régulation.
 6
7
6
7
1
/
7
100%