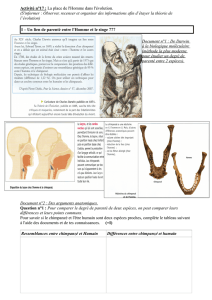
Parenté entre êtres vivants actuels et fossiles – phylogenèse

1
Parenté entre êtres vivants actuels et fossiles –
phylogenèse - évolution.
Introduction.
Définition des termes du titre puis rappels de collège et de lycée.
Reconstitution de l’histoire évolutive de différentes lignées depuis leur
divergence à partir d’un ancêtre commun
L’idée d’évolution peut être illustrée à l’aide de plusieurs exemples
vus dans les classes antérieures :
Pré-requis :
En sixième : les élèves apprennent à classer les êtres vivants à partir
de comparaison de caractères.
En quatrième : la notion d’espèce et d’évolution des êtres vivants et de
leur milieu.
En seconde : Origine commune des espèces vivantes confirmant la
notion d’évolution, mise en évidence au niveau de la cellule, de
l’embryon et de la molécule.
En 1er S : relation entre gène et protéine, universalité du code
génétique et des modalités d’expression des gènes à renforcer l’idée
de l’origine commune des êtres vivants.
Problématique générale : Comment peut-on établir le degré de
parentés entre les êtres vivants ?
1. La recherche des liens de parenté chez les êtres
vivants.
a) La parenté entre les êtres vivants à partir des données
morphologiques.
Caractères
Age du plus
ancien fossile
possédant ce
caractère (en
Groupes
Exemple
d’organisme
appartenant à
ce groupe

2
Ma)
Poils
220
Mammifère
Rat
Fort
plissement du
cortex cérébral
23
Hominoïde
Orang-outan
Vertèbres
470
Vertébrés
Lamproie
Noyau à
l’intérieur des
cellules
1000
Eucaryote
Chêne
Amnios de
l’embryon
340
Amniote
Pigeon
Pouce
opposable
55
Primate
Ouistiti
Fusion
prénatale de
certains os du
poignet
10
Hominidé
Gorille
Membres pairs
munis de
doigts
370
Tétrapode
Grenouille
Soudure de
certains os du
palais
7
homininé
Chimpanzé
Différents caractères permettant de justifier la position
phylogénétique de l’homme actuellement.
A l’aide des explications de la page 291, représentez l’arbre
phylogénétique reliant l’Homme et les autres organismes cités.
Comment a-t-on classé toutes ces espèces différentes ?
Nous avons pu classer ces différents groupes grâce au partage d’un
plus ou moins grand nombre de caractères dits homologues (structures
ayant une même organisation d’ensemble, une même origine
embryologique et les mêmes connexions avec l’organisme).

3
Comment établit-on des relations de parenté ? Quand peut-on
dire que deux groupes sont particulièrement proches ?
Des relations de parenté sont établies après l’étude des caractères
homologues, pour lesquels sont définis un état ancestral et un état
dérivé issu de la transformation de cet état primitif.
Deux êtres vivants qui possèdent un nombre élevé de caractères dans
le même état dérivé ont une parenté étroite.
A quoi correspond un « nœud » dans un arbre phylogénétique ?
Le nœud correspond à l’ancêtre commun entre plusieurs êtres vivants.
Cet ancêtre présente souvent l’apparition d’un nouveau caractère. Ce
caractère tel qu’il est apparu est inconnu ; on ne connaît que l’état
dérivé (exemple : l’apparition des membres pairs munis de doigts, il
en existe beaucoup de différentes qui sont résultats de l’évolution).
OU
TP PHYLOGENE.
Bilan :
L’établissement d’une relation de parenté est possible en
comparant les différents états d’un même caractère. Les
caractères choisis sont dits « homologues » car elles doivent
occuper une position identique à l’intérieur d’un plan
d’organisation partagé par plusieurs individus. Ces caractères
peuvent se présenter sous différents aspects (embryonnaire,
morphologique ou anatomique)
Les modifications que les structures homologues ont subies au
cours de l’évolution définissent des états de caractères nouveaux,
dérivés d’états de caractères ancestraux (passage d’un caractère
dit primitif à un caractère dit dérivé). L’état dérivé est celui qui
est apparu le plus récemment au cours des temps géologiques.

4
Chaque branche entre deux nœuds doit être justifiée par au moins
une innovation évolutive (ou état dérivé, ou évolué, d’un
caractère).
Chaque nœud d’un arbre phylogénétique correspond au dernier
ancêtre commun. Il restera toujours hypothétique, et on ne
pourra que reconstituer quelques-unes de ses caractéristiques : il
possède toutes les innovations évolutives acquises par ses
précédents ancêtres.
L’évolution des caractères homologues nous renseignent ainsi sur
la plus ou moins grande parenté de certains organismes : le
partage d’états dérivés de certains caractères témoigne d’une plus
étroite parenté que le partage d’états ancestraux de ces mêmes
ancêtres.
L’établissement d’une relation de parenté est possible en comparant
les différents états d’un même caractère (embryonnaire (document 1
page 20), morphologique (document 2 page 20) ou anatomique
(document 1,2 et 3 page 22 et 23)). Les caractères choisis peut se
présenter sous différents aspects, amenant l’idée d’évolution chez les
descendants d’un ancêtre possédant ce caractère (document 2 page 22
exemple des dents de proboscidiens). Ces caractères sont dits
homologues.
Des structures homologues occupent une position identique à
l’intérieur d’un plan d’organisation partagé par plusieurs individus
(document 1 page 20-21 et document 3 page 23) : ce sont elles que
l’on peut comparer puisqu’elles dérivent d’une même structure
ancestrale.
Les modifications que les structures homologues ont subi au cours de
l’évolution définissent des états de caractères nouveaux, dérivés
d’états de caractères ancestraux (passage d’un caractère dit primitif à
un caractère dit dérivé). (Document 3 page 23)
L’évolution des caractères homologues nous renseignent ainsi sur la
plus ou moins grande parenté de certains organismes : le partage

5
d’états dérivés de certains caractères témoigne d’une plus étroite
parenté que le partage d’états ancestraux de ces mêmes ancêtres.
b) Le traitement et l’apport des données moléculaires
pour l’établissement d’un arbre phylogénétique.
TP sur Phylogène et Anagène.
Le cas de la molécule CDC2.
Hérédité des modalités de la division cellulaire et gène CDC2
Dans un milieu favorable et une température de 35°, la levure
Schizosaccharomyces Pombe se divise toutes les 3 heures environ.
L'entrée en division en fonction de la taille est une caractéristique
héréditaire.
hapl.gif
Parmi les gènes en jeu, le gène CDC2 joue un rôle majeur : il code
pour une protéine qui exerce un contrôle sur deux points cruciaux du
cycle cellulaire, l'un est l'entrée en phase S, et l'autre est l'entrée en
mitose. Le point de contrôle le plus important dans le cycle cellulaire
de la levure est le point d'entrée en mitose. Le gène CDC2 est un
composant décisif pour conduire les cellules en mitose ; lorsqu'il est
défectueux, la mitose ne peut pas se produire. Les travaux sur le gène
CDC2 et sur les autres gènes régulateurs du cycle cellulaire ont valu à
P. Nurse, L Harwell et T. Hunt le prix Nobel de médecine 2001.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
1
/
28
100%