Chapitre 11

Chapitre 11
La régulation de l’expression des gènes chez les bactéries et leurs virus
Une représentation du complexe répresseur lac-ADN, telle
qu’elle a été déterminée par crystallographie aux rayons X

Régulation de l’activité des gènes
Procaryote
Eucaryote Phénomène de différentiation
Les concentrations de protéines peuvent être régulées par des
mécanismes qui opèrent lors de la transcription ou de la traduction.

La régulation des gènes chez les procaryotes
La régulation de la transcription peut être placée sous contrôle négatif ou
positif. Sous contrôle négatif, l’expression génique aura lieu sauf si elle est
inhibée par une molécule régulatrice (répresseur). À l’opposé, sous contrôle
positif, la transcription à lieu seulement si une molécule régulatrice
(activateur) stimule directement la production d’ARNm.

Effecteurs allostériques
Les activateurs et les répresseurs agissent comme des commutateurs
génétiques. Ces protéines possèdent deux sites importants: un domaine de
liaison à l’ADN et un site allostérique. Le site allostérique interagit avec de
petites molécules appelées effecteurs allostériques, modifiant ainsi la capacité
des activateurs et répresseurs à se lier à l’ADN.

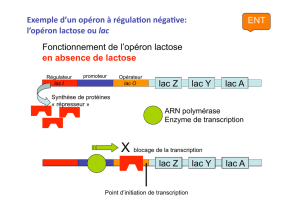
L’exemple du métabolisme du lactose
Un minimum de deux enzymes est nécessaire au métabolisme du lactose:
1- Une β-galactosidase: Scinde le lactose en deux molécules distinctes de
galactose et de glucose.
2- Une perméase: Nécessaire au transport du lactose à l’intérieur de la cellule.
 6
7
8
9
10
11
12
13
14
15
16
17
6
7
8
9
10
11
12
13
14
15
16
17
1
/
17
100%