pdf file - FR

Unité d'ouverture
INTELLIGENCE ARTIFICIELLE
Partie 2: Réseaux de neurones biologiques
Unité d'ouverture
Systèmes Adaptatifs à Base d ’Apprentissage
(SBA- Ingénieurs I5)
Tarik AL ANI, Département Informatique
ESIEE-Paris
E-mail : [email protected]r
Url: http://www.esiee.fr/~alanit

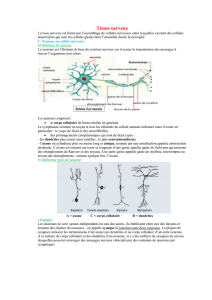
LE CERVEAU HUMAIN
06/11/2011 1

RESEAUX DE NEURONES BIOLOGIQUESRESEAUX DE NEURONES BIOLOGIQUES
Pèse environs 1500 grammes et contient
trois couches successives :
le cerveau reptilien, siège des réflexes
instinctifs,
06/11/2011 2
l’hippocampe, siège de l ’émotivité et de la
sensibilité,
le cortex, spécifique aux mammifères et
siège du raisonnement et de la pensée.

Ces couches successives contiennent deux
types du cellules :
les gliales constituent le tissu nourricier et
la structure du cerveau et représentent
environs
99
.
99
%
du
volume
du
cerveau
.
06/11/2011 3
environs
99
.
99
%
du
volume
du
cerveau
.
les neurones dont le nombre, estimé à un
nombre variant entre 10-100 milliards,
forment un réseau interconnecté complexe.

Trois types de neurones :
1. Les neurones sensitifs
C ’est le long de ces neurones que voyagent
les
signaux
nerveux
.
06/11/2011 4
les
signaux
nerveux
.
Exemple :
Signaux qui proviennent de récepteurs de la
peau et qui sont transférés vers le système
nerveux central.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
1
/
45
100%