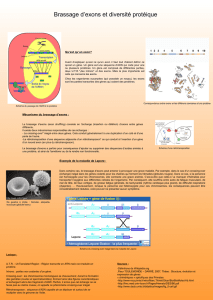
Génétique et particules d`information redistribution des exons au

n°
G
·
Figure
51
1
/
1
100%