SYNTHESE ET ACTIVITE ANTIMICROBIENNE DE

Rev. Microbiol. Ind. San et Environn. Vol 7, N°1, p : 1-29 Chennaoui et al., 2013
1
SURVIE DES BACTERIES ISSUES DU REJET D’ABATTOIR
DANS LES EAUX DE LA PLAGE D’EL JADIDA (MAROC)
M. CHENNAOUI1*, H. AASSILA2, Y. FARID1, A. EL KOURI1, M.
MOUNTADAR2 et O. ASSOBHEI1
1- Laboratoire de Biotechnologie Marine et de l’Environnement (Biomare).
2- Unité de Chimie Analytique et Génie de l’Environnement.
Faculté des Sciences, Université Chouaib Doukkali, BP : 20, El Jadida 24000 (Maroc).
Pour correspondance : [email protected]
RESUME
La présente étude sur la survie des bactéries issues du rejet d’abattoir dans l’eau
de mer a été réalisée au laboratoire et insitu. Les facteurs abiotiques étudiés au
laboratoire sont le rayonnement solaire, la température et la salinité. Pour évaluer l’effet
des facteurs biotiques et abiotiques sur la survie des bactéries dans les conditions
naturelles, des chambres de diffusion ont été utilisées. Les résultats mettent en relief
l'importance de ces paramètres de stress dans l’inactivation des bactéries. Les
pourcentages de dormance peuvent atteindre 98% à forte salinité. Les résultats obtenus
des essais in situ montrent une réduction des effectifs bactériens plus importante que
lors de l’étude menée au laboratoire. L'étude comparative des techniques de comptage
utilisées montre que les facteurs abiotiques et biotiques ont un effet bactéricide ou
bactériostatique, ce qui entraîne la transformation des bactéries à un état viable non
cultivable leur permettant de conserver leur caractère pathogène.
Mots clés : Abattoir, survie, bactérie.

Rev. Microbiol. Ind. San et Environn. Vol 7, N°1, p : 1-29 Chennaoui et al., 2013
2
ABSTRACT
The present study on the survival of bacteria from the rejection of an abattoir in
seawater was carried out in the laboratory and in situ. Abiotic factors studied in the
laboratory are solar radiation, temperature and salinity. To evaluate the effect of biotic
and abiotic factors on the survival of bacteria in natural conditions, diffusion chambers
were used. The results highlight the importance of these stress parameters in the
inactivation of bacteria. The percentages of dormancy can reach 98% to high salinity.
The results of in situ tests show a reduction in bacterial numbers greater than during the
study in the laboratory. The comparative study of counting techniques used shows that
the abiotic and biotic factors have a bactericidal or bacteriostatic effect, which causes
the transformation of bacteria viable non-culturable state allowing them to retain their
pathogenicity.
Keywords: Abattoir, survival, bacteria.
INTRODUCTION
La plage de la ville d’El jadida, située sur la côte atlantique marocaine est
caractérisée pour son importante activité touristique estivale et par ses richesses
aquacoles. Cependant, cette plage est soumise à une forte pollution par les eaux usées
parmi lesquelles les rejets d’abattoir. Ces derniers contiennent de nombreux organismes
pathogènes (ex : Salmonelles, virus de l'hépatite A...), représentant une menace pour la
santé humaine et l’écosystème marin (Lamghari et Assobhei 2005, Chennaoui et al.,
2006).
Il est bien connu que lorsque les bactéries issues de ses rejets arrivent dans
l’environnement côtier, leur devenir dépend à la fois de processus hydrodynamiques,
biotiques (liés à l’action des êtres vivants) et physiologiques (action sur les fonctions

Rev. Microbiol. Ind. San et Environn. Vol 7, N°1, p : 1-29 Chennaoui et al., 2013
3
propres de la cellule) (Troussellier et al., 1998).
Jusqu'au milieu des années 70, il semblait bien établi que les bactéries étaient
vouées soit à une disparition plus ou moins rapide , soit à une altération de leur état
physiologique sous l'influence du pouvoir "auto-épurateur" du milieu marin :
température basse, salinité élevée, irradiation solaire intense, carence nutritionnelle,
présence de substances toxiques des eaux usées (détergents, métaux lourds), activité de
nombreux micro-et macro-prédateurs, inhibition ou lyse par des substances
antibiotiques produites par les microorganismes marins (algues, champignons,
bactéries) (Aubert et al., 1981).
Ce concept de "mortalité" bactérienne a rapidement évolué au cours des quinze
dernières années. En effet, la revivification des bactéries telluriques d'origine fécale,
dans certains milieux particulièrement agressifs (eaux chlorées, eaux froides ou
chaudes, eaux salées, etc...), éventuellement réversible par l'utilisation de techniques de
dénombrement dites de "reviviscence" (Garry 2000).
Nous étudierons successivement le comportement des bactéries dans le milieu
marin, en l’exposant, au laboratoire et in situ, aux paramètres biotiques et abiotiques du
milieu qui contrôlent leur survie. Ainsi, nous avons choisi quatre espèces de bactéries
dont les abondances sont élevées (E. coli, C. perfringens, St. aureus et Streptocoques.
fécaux).
MATERIEL ET METHODES
Description du site d’étude
Le système côtier de la ville d’El Jadida constitue l’assise d’une activité
multiforme (pêche, ramassage des algues, exploitation des sables, tourisme
balnéaire…). Malheureusement, tous les rejets de la ville, dont celui de l’abattoir
déversent en mer ce qui constitue une menace sérieuse pour cet environnement marin et

Rev. Microbiol. Ind. San et Environn. Vol 7, N°1, p : 1-29 Chennaoui et al., 2013
4
notamment le littoral d’El Jadida (figure 1).
Figure 1 : Localisation de la ville d’El Jadida et de son abattoir municipal
Préparation des suspensions bactériennes
Souches
Les souches utilisées dans le présent travail sont des collections du laboratoire
de Biotechnologie marine et de l’environnement (Biomare) d’El jadida. Il s’agit de C.
perfringens, E.coli, des streptocoques fécaux et Streptococcus aureus.
Milieux de culture
Les milieux de culture utilisés ainsi que la température d’incubation pour les
différents microorganismes étudiés sont Lactose Sulfite (LS) à 37°C pour C.
perfringens, Lysogeny broth (LB) à 44°C pour E. coli, le milieu Ethyl Violet Azide

Rev. Microbiol. Ind. San et Environn. Vol 7, N°1, p : 1-29 Chennaoui et al., 2013
5
(EVA) à 37°C pour les streptocoques fécaux et le milieu chapman à 37°C pour S.
aureus
Cultures
Les cellules en phase exponentielle de croissance sont récoltées par
centrifugation (3000 g pendant 15 min). Les culots sont lavés deux fois avec une
solution de NaCl à 90/00 et mis en suspension dans la même solution. A partir de ces
suspensions, nous avons ensemencé les milieux de culture à raison de 10% dans des
erlens et les chambres de diffusion contenant une solution d’eau de mer pour étudier
l’effet des facteurs biotiques et l’effet des facteurs abiotiques sur leur survie. Les
évolutions des abondances bactériennes sont suivies toutes les 24 heures pendant 10
jours par dénombrement indirect sur milieux de culture spécifiques et par
dénombrement direct en utilisant la technique d’épifluorescence au DAPI (4’,6-
diamidino-2-phenylindole, Sigma).
Comptage des bactéries étudiées
1- Comptage indirect
Les unités forming colony (UFC) des différentes espèces bactériennes ont été réalisées
dans les milieux de culture solide résumés dans le tableau I.
Tableau I : milieux utilisés pour le dénombrement des différentes espèces.
Espèce
Milieu sélectif
Incubation
Milieu non sélectif
Incubation
E.coli
EMB Biokar
44°C/24h
TSA + 6g/l d’E.L
37°C/24h
C. perfringens
LS
46°C/8h
TSA + 6g/l d’E.L
37°C/24h
S. aureus
MSA
37°C/24h
TSA + 6g/l d’E.L
37°C/24h
Streptocoques
fécaux
SB
37°C/24h
TSA + 6g/l d’E.L
37°C/24h
E.L Extrait de levure
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
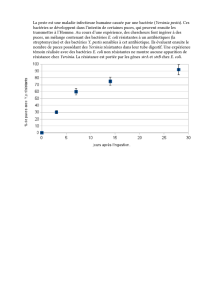
1
/
29
100%