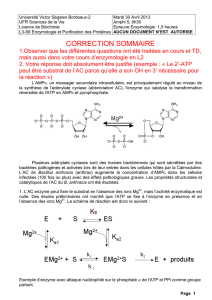
Les CREB à la CREM - iPubli

506
E,
•••
1'
• •• BRÈVES •••
° 91
1
/
1
100%