cours 1S

- 1 cours 1S-
© N. Bouchaud
SVT CLASSE DE 1ère S ANNEE 2006/2007 LYCEE Descartes
PARTIE I : SCIENCES DE LA VIE.
DES PHENOTYPES A DIFFERENTS NIVEAUX D’ORGANISATION DU VIVANT
Ch1 DU GENOTYPE AU PHENOTYPE, RELATIONS AVEC L’ENVIRONNEMENT
Introduction.
L’ADN est le support universel de l’information génétique. Notre « apparence » dépend donc de lui.
Quelles sont les relations entre nos gènes et notre apparence ? D’autres facteurs externes peuvent-ils
intervenir ?
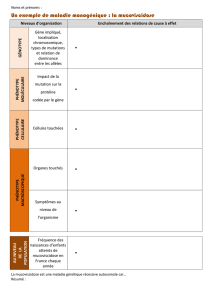
1 Le phénotype se définit à différentes échelles.
Le phénotype [ ] est l’ensemble des caractères observables d’un individu. Il peut se définir
aux différentes échelles d’organisation du vivant allant de l’organisme à la molécule. Dans le cas de la
drépanocytose, le niveau macroscopique (organisme) correspond aux symptômes de la maladie (ou
au fait qu’on soit sain), le niveau cellulaire aux altérations cellulaires (ici en faucille ou biconcave) et
enfin le niveau moléculaire à la protéine déficiente ou non (hémoglobine). Les différents niveaux
sont liés entre eux (la molécule change la forme de la cellule et entraîne les symptômes) ! Noter ici que
l’environnement intervient pour déclencher les symptômes.
Voir l’exemple de la drépanocytose.
2 La réalisation du phénotype nécessite l’intervention des protéines.
Dans l’exemple de la drépanocytose, c’est l’allèle morbide qui entraîne la maladie, mais
indirectement par la synthèse d’une protéine déficiente.
Une protéine (on parle aussi de polypeptide) est une macromolécule constituée d’une
séquence définie d’acides aminés. Elle est fabriquée par un gène. Une protéine a une fonction précise
(transport d’O2 dans le cas de l’hémoglobine). Il existe un répertoire de 20 acides aminés différents.
Leur enchaînement par des liaisons peptidiques correspond à la structure primaire des protéines.
Les protéines sont des molécules 3D : les acides aminés interagissent entre eux (liaisons
faibles par exemple) et conduisent à la structure secondo-tertiaire. Enfin l’association entre plusieurs
chaînes polypeptidiques définit la structure quaternaire (exemple de l’hémoglobine).
Voir le schéma page 162.
Les phénotypes alternatifs (variations d’un même caractère entre différents individus)
sont donc dus à des différences dans les séquences d’acides aminés des protéines.
Voir avec la drépanocytose.
3 Les enzymes sont des protéines essentielles dans la réalisation du phénotype.
3.1 Les enzymes accélèrent les réactions chimiques
Les protéines enzymatiques sont des catalyseurs biologiques (ou biocatalyseurs) c’est-à-dire
qu’elles accélèrent les réactions chimiques (ou permettent leur réalisation) sans subir elles-mêmes de
transformations. Cela signifie qu’après avoir agit, l’enzyme reste fonctionnelle.
Le schéma de la réaction est le suivant :
E + S ES E + P
A noter que les enzymes agissent surtout à faible concentration.
3.2 Les enzymes possèdent une double spécificité.
Les enzymes présentent :
- une spécificité de substrat puisqu’elles n’agissent que sur une seule molécule spécifique.
- une spécificité d’action car elles ne catalysent qu’un seul type de réaction chimique.
Cependant, une molécule peut être le substrat d’une multitude d’enzymes. L’enzyme agit sur le
substrat en formant ce qui s’appelle un complexe enzyme. Le substrat interagit avec l’enzyme au
niveau de son site actif (région en forme de « creux »).
Le site actif comprend :
- des acides aminés de liaison. Ils maintiennent le substrat par des liaisons faibles et
permettent la spécificité de substrat ;
- des acides aminés catalytiques qui effectuent la réaction chimique et qui permettent
la spécificité d’action. Ce complexe enzyme – substrat est temporaire.
Inclure le graphique du Belin page 177 (vi = f([S]).
La courbe a la forme d’une hyperbole. La vi augmente très rapidement et atteint un maximum quand toutes les
enzymes sont engagées dans la catalyse : il y a donc saturation.
Formation du complexe enzyme – substrat (schématisation) : Nathan p 43.

- 2 cours 1S-
© N. Bouchaud
3.3 L’activité enzymatique dépend des conditions du milieu.
L’activité des enzymes contribue à la réalisation du phénotype.
Les propriétés des enzymes dépendent de leur structure spatiale (cf. 2.1). Des changements de
structure primaire (séquence d’acides aminés) peuvent modifier la structure spatiale de la molécule :
l’enzyme peut devenir non fonctionnelle (modification des liaisons chimiques intra-chaîne d’où une
forme différente). Le phénotype macroscopique ou cellulaire est alors altéré.
Les conditions de l’environnement modifient également leur activité (pH et température
incompatibles avec l’activité des protéines…).
- Température : une basse température diminue l’agitation moléculaire alors qu’une forte
température dénature la protéine.
- pH : des pH trop bas ou trop élevés dénaturent la protéine.
- Attention : pH et température sont propres à chaque enzyme.
4 Du gène aux protéines.
Le phénotype dépend de l’expression des protéines. Les protéines, enchaînement d’AA, sont
fabriquées à partir des gènes. On trouve environ 30 000 gènes dans le génome humain. Comment
passe-t-on d’un gène à une protéine ?
4.1 On fabrique une copie du gène…
La transcription correspond à la synthèse d’un acide nucléique proche de l’ADN : c’est
l’ARNm (pour acide ribonucléique messager). Contrairement à l’ADN, l’ARNm est une molécule qui
possède un simple brin complémentaire du brin transcrit de l’ADN (qui sert de matrice).
L’appariement des nucléotides entre le brin d’ADN transcrit et l’ARNm est le même qu’entre les deux
brins d’ADN (G avec C, T avec A, mais A avec U puisque l’uracile remplace la thymine dans l’ARN).
Une fois synthétisé, l’ARNm quitte le noyau par les pores nucléaires et passe dans le
cytoplasme des cellules eucaryotes.
C’est une enzyme qui réalise la transcription : l’ARN polymérase. Elle se fixe, ouvre et
déroule la molécule d’ADN puis incorpore des nucléotides complémentaires (= elle polymérise les
nucléotides).
NB : L’ARN est une molécule à courte durée de vie ;
De multiples copies sont produites (amplification).
Il n’y a pas qu’un seul brin transcrit dans l’ADN (brins antiparallèles).
4.2 … Avant de synthétiser la protéine.
La traduction permet la synthèse de chaînes polypeptidiques à partir de l’ARNm et d’un
répertoire de 20 AA. Elle se déroule dans le cytoplasme des cellules. La séquence des acides
aminés est codée par celle des nucléotides de l’ARN messager suivant un système de
correspondance, le code génétique. La correspondance est de trois nucléotides de l’ARN (on parle de
codon) pour un acide aminé. Ce code génétique est universel (procaryotes et eucaryotes possèdent le
même) et dégénéré (plusieurs codons peuvent correspondre au même acide aminé) donc redondant.
La traduction débute toujours au codon d’initiation (qui correspond à la méthionine) et s’arrête
au codon stop (trois possibilités différentes) qui ne correspond à aucun AA.
On trouve plusieurs étapes dans la traduction :
- l’initiation est caractérisée par la fixation d’un ribosome sur un codon de l’ARNm
(AUG)
- l’élongation du polypeptide par déplacement du ribosome sur l’ARNm. Réalisation
de la liaison peptidique.
- la terminaison, provoquée par l’arrivée du ribosome sur un codon stop. Le ribosome
se dissocie de l’ARNm et libère le polypeptide.
NB : la molécule qui joue le rôle d’adaptateur entre les AA et les codons est l’ARNt.
Là encore, notion d’amplification.
S’inspirer du schéma bilan p60 et 61 (Hatier).
5 Les relations entre gènes, phénotypes et environnement sont complexes.
On veut montrer qu’à un génotype précis, ne correspond pas un seul phénotype.
Génotype : ensemble des allèles d’un être vivant. On envisage souvent le génotype impliqué dans
l’établissement d’un phénotype ; il se matérialise alors par l’écriture des deux allèles du gène
considéré.

- 3 cours 1S-
© N. Bouchaud
- Première complication :
Il y a deux copies d’un gène par cellule (les allèles). Les copies peuvent être identiques
(homozygote) ou différentes (hétérozygote). On prend l’exemple de la drépanocytose. (HbS/HbS =
malade ; HbA/HbA et HbA/HbS = sain).
Ici on a deux génotypes possibles pour un phénotype macroscopique sain car HbS est
récessif. En général, la présence d’une seule copie fonctionnelle du gène assure un phénotype normal
(allèle HbS récessif). PS : HbS s’exprime mais la présence d’HbA l’empêche de polymériser.
- Deuxième complication.
Un phénotype macroscopique donné peut résulter de l’expression de plusieurs gènes
(exemple, les groupes sanguins : on parle de chaîne de biosynthèse). La mutation de l’un seulement
de ces gènes peut altérer ce phénotype. Un même phénotype macroscopique peut donc
correspondre à plusieurs génotypes.
Voir la chaîne de biosynthèse correspondant aux groupes sanguins.
Exemple des groupes sanguins : le gène du groupe sanguin présente 3 allèles : A, B et O (PS il y a
même de multiples versions de A…). Donc plusieurs génotypes A conduisent au même phénotype.
Même chose pour la PCU (300 allèles).
- Troisième complication.
Chez un individu donné, l’effet des allèles d’un gène va dépendre également de
l’environnement : un même génotype peut donner des phénotypes macroscopiques différents en
fonction des conditions environnementales (ce n’est pas le cas pour tous les gènes !). (xeroderma,
couleur des fleurs, PCU, drépanocytose).
Conclusion.
Le génotype est responsable de l’élaboration du phénotype à travers la synthèse protéique,
dont les enzymes en sont un exemple.
Un même phénotype peut correspondre à plusieurs génotypes, mais phénotype peut résulter de
l’action conjointe du génotype et de l’environnement.
Ch2. LA MORPHOGENESE VEGETALE ET L’ETABLISSEMENT DU PHENOTYPE
Introduction.
Le phénotype d’un individu est le résultat des interactions entre l’expression de son génotype
(diversité allélique) et d’un contrôle par l’environnement (diversité des facteurs externes). De ce fait,
il existe une diversité phénotypique individuelle.
La morphogenèse végétale constitue un exemple de cette diversité et de ces interactions.
Morphogenèse : ensemble des mécanismes qui assurent la mise en place et la différenciation
des organes lors du développement des organismes animaux et végétaux.
Les organes des végétaux sont les tiges, les feuilles, les racines, les fleurs…
1 Les végétaux présentent une grande diversité de formes.
Tous les végétaux ont la même organisation générale :racines pour l’ancrage dans le sol, prélèvement
d’eau et de sels minéraux, tige aérienne avec feuilles permettant la photosynthèse.
On distingue le port (allure générale d’une plante) arborescent, buissonnant et herbacé.
Les bourgeons (organe végétatif comprenant un méristème apical, des ébauches de feuilles et
bourgeons axillaires, responsable de la construction de la partie aérienne de la plante) assurent la mise
en place des parties aériennes d’une plante.
1.1 La morphologie des végétaux est liée à l’expression des gènes.
Au sein d’une même espèce végétale, les individus présentent des caractères morphologiques
relativement constants et transmis d’une génération à la suivante : la morphologie d’un végétal dépend
donc en partie des caractéristiques génétiques de l’espèce (donc de son génotype). Page 244.
1.2 L’expression du génotype est modulée par l’environnement.

- 4 cours 1S-
© N. Bouchaud
En fonction de leur environnement, des individus d’une même espèce peuvent toutefois
posséder une morphologie différente. Les végétaux, contrairement aux animaux ne peuvent se
déplacer : ils doivent s’adapter aux conditions environnementales. Ils possèdent une certaine plasticité
de forme. Cette plasticité est permise par le fait que les végétaux grandissent toute leur vie (croissance
indéfinie). Elle est influencée par :
- des facteurs abiotiques : température, vent, lumière (page 246, 248)
- des facteurs biotiques : champignons, bactéries, compétition entre espèces (page 247).
Cette morphologie est transmissible (adaptation) ou non à la descendance (accomodation).
Note : l’homme intervient aussi en taillant les végétaux (bonsaï).
1.3 Les contraintes du milieu peuvent « sélectionner » des morphologies.
Des réponses morphologiques semblables peuvent être obtenues avec des végétaux d’espèces
différentes placés dans un même environnement : il s’agit d’une convergence morphologique. C’est
le cas des espèces de milieux arides comme les plantes succulentes : Cactées en Amérique et
Euphorbes en Afrique. Leurs tiges sont sphériques ou cylindriques avec des cellules gorgées d’eau,
des pertes d’eau réduites par forme sphérique, des feuilles réduites (ou aiguillons). C’est une
adaptation génétiquement acquise. Livre page 249.
2 La morphogenèse associe la division et la croissance cellulaire.
Un végétal possède toute sa vie des cellules embryonnaires : elles se trouvent dans les
méristèmes (localisés à l’extrémité des racines ou dans les bourgeons).
La croissance est l’augmentation au cours du temps de caractères quantitatifs comme la masse ou la
taille.
2.1 La croissance végétale dépend de l’activité des méristèmes.
Les divisions cellulaires (mitoses) ont lieu dans les méristèmes. Ils sont situés aux extrémités
racinaires mais également au niveau des tiges de la plante, dans les bourgeons (bourgeon apical et
axillaire). Ils sont responsables de la morphogenèse végétale. La morphogenèse comprend une phase
de multiplication et de croissance cellulaires puis de différenciation des organes (ou organogenèse).
Les méristèmes contiennent des cellules méristématiques, cellules embryonnaires non
différenciées : petites vacuoles, gros noyau, petit cytoplasme.
2.2 Les cellules méristèmatiques ont deux destinées différentes.
Les mitoses produisent :
- Des cellules qui vont ensuite s’allonger et se différencier (en arrière des méristèmes)
et qui vont participer à la croissance et à la structuration de l’organisme (feuilles,
tiges…) ;
- Des cellules qui restent indifférenciées et qui vont à leur tour constituer des
méristèmes. Bilan : schéma bilan (Bordas) et TP1.
3 La mitose est un processus commun aux cellules eucaryotes.
Une cellule subit un ensemble de changements entre deux divisions de mitose : c’est le cycle
cellulaire. Ce dernier comprend l’interphase et la mitose.
3.1 L’ADN se réplique au cours de l’interphase.
Pendant l’interphase, la cellule effectue ses synthèses. L’interphase est découpée en trois
phases.
- La phase G1 qui suit la division cellulaire précédente (activité métabolique).
- La phase S qui correspond au doublement de la quantité d’ADN dans le noyau d’une
cellule. L’ADN se réplique selon un mécanisme semi-conservatif, fondé sur la
complémentarité des bases (A avec T ; G avec C). Cette réplication nécessite l’ouverture
de la molécule d’ADN (et donc sa détorsion) et la synthèse d’un nouveau brin à partir du
précédent. C’est un complexe enzymatique, l’ADN polymérase, qui catalyse la
polymérisation des nucléotides. La réplication se produit en plusieurs points de la
molécule d’ADN (yeux de réplication) afin d’accélérer le processus.
- La phase G2 qui suit la réplication de l’ADN. Pendant cette phase, la cellule se prépare à
entrer en mitose.
Bilan : TP3 et schéma bilan (Bordas).
3.2 Les structures cellulaires se modifient lors de la mitose.
L’enveloppe nucléaire, donc le noyau, disparaît, la chromatine se condense en chromosomes à
deux chromatides sœurs (issues du doublement de la quantité d’ADN pendant la phase S), réunies au

- 5 cours 1S-
© N. Bouchaud
niveau du centromère (prophase). Ils se regroupent à l’équateur de la cellule formant la « plaque
équatoriale » (métaphase) avant de se scinder en deux au niveau du centromère et de migrer à chaque
pôle de la cellule (anaphase) et de se décondenser en télophase, ultime phase qui marque la fin de la
mitose et qui s’achève par la cytodiérèse (formation de la paroi et séparation des deux cellules filles).
Les chromosomes sont « tirés » vers les pôles de la cellule par des structures particulières : le
cytosquelette (système de filaments protéiques qui permet des mouvements cellulaires contrôlés).
Bilan : schémas du TP2 et Bordas.
3.3 Le patrimoine génétique est conservé lors de la mitose.
Il y a conservation de l’information génétique lors de la mitose : chaque cellule fille hérite
d’une information génétique identique à celle de la cellule mère (reproduction à l’identique). La seule
différence concerne la quantité d’IG qui est divisée par deux (chaque chromosome d’une paire n’a
plus qu’une chromatide). Ainsi,le doublement de la quantité d’ADN et la mitose permettent la
conservation de l’IG au cours des générations.
4 La croissance cellulaire est contrôlée par une hormone végétale, l’auxine.
4.1 La paroi végétale des cellules en croissance est extensible.
La paroi des cellules végétales en extension est essentiellement constituée de polysaccharides
(polymères biologiques dont les unités sont des molécules de sucre), dont la cellulose organisée en
fibre (polymères linéaires de glucose), les pectines et les hémicelluloses (sorte de gel hydraté). Des
protéines viennent s’y ajouter. Tous ces éléments sont reliés par des liaisons faibles et covalentes. La
paroi est donc un matériau composite (formé d’éléments très différents).
Les molécules de cellulose forment des chaînes parallèles reliées par des liaisons faibles et qui
sont réunies en microfibrilles. Les microfibrilles sont dans une matrice plus ou moins gélatineuse
formée par la pectine, ce qui permet le glissement des fibrilles de cellulose et rendant la paroi
extensible lorsqu’elle est jeune. Schéma TP et page 292.
4.2 La croissance cellulaire repose sur la turgescence et la plasticité pariétale.
La paroi des cellules jeunes possède une certaine plasticité ; c’est-à-dire qu’elle peut s’étirer et
permettre la croissance cellulaire (déformation irréversible contrairement à l’élasticité des cellules
adultes). Les différentes fibres constitutives de la paroi glissent les unes par rapport aux autres par
rupture des liaisons hydrogène.
Ce glissement se produit lorsque la vacuole végétale est turgescente : elle provoque une
pression, ou pression de turgescence, qui s’exerce sur la paroi, ce qui lui permet de s’étirer. La vacuole
étant hypertonique (plus concentrée que le milieu extérieur à cause de la présence d’ions, d’acides
aminés, de saccharose…), l’eau à tendance à y entrer par osmose (passage d’eau vers le milieu le plus
concentré à travers une membrane semi-perméable). L’état naturel d’une cellule est la turgescence.
N.B. : une cellule en état de stress hydrique est une cellule plasmolysée.
La paroi empêche la cellule d’éclater.
INTRODUIRE LES TERMES : hypotonique, hypertonique, turgescence et plasmolyse.
4.3 L’auxine contrôle la croissance cellulaire.
Une hormone est un messager chimique entre deux cellules. Une hormone végétale doit être
fabriquée par la plante elle-même (et non pas absorbée dans le milieu de vie), agir à très faible
concentration, véhiculer une information à des cellules cibles éloignées ou non, sensibles à son action
(cellules pourvues de récepteurs), dont elle modifie le fonctionnement. Elle possède de multiples
actions dans le végétal.
Ajouter le schéma.
L’auxine est synthétisée par l’apex des coléoptiles de Graminées, les méristèmes et les jeunes
feuilles des bourgeons terminaux des plantes en général. Elle dérive de l’acide aminé tryptophane. Il
existe gradient d’auxine dans la plante : sa concentration décroît des apex des tiges vers les racines.
Elle se fixe sur des récepteurs spécifiques ancrés dans la membrane plasmique des cellules.
Elle possède une double action dans la croissance :
- une action à court terme sur la plasticité pariétale. L’auxine stimule la rupture des
liaisons hydrogène entre les divers constituants de la paroi et donc l’extensibilité de la
paroi par acidification (expulsion de protons) ce qui permet un coulissage entre molécules.
Par la suite, cela active des enzymes qui hydrolysent les liaisons fortes.
- une action à plus long terme sur l’expression de gènes qui participent aux divers
événements du métabolisme nécessaires à la croissance (stimulation de la synthèse
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
1
/
20
100%