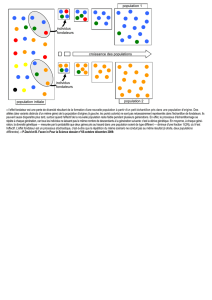
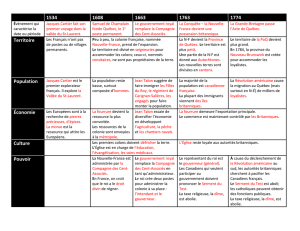
étude comparative des caracteristiques généalogiques

CATHY CAUVIER
ÉTUDE COMPARATIVE DES CARACTERISTIQUES
GÉNÉALOGIQUES ET GÉNÉTIQUES DE QUATRE
POPULATIONS FONDATRICES DE LA GASPESIE
Mémoire présenté
à la Faculté des études supérieures de l'Université Laval
comme exigence partielle du programme de maîtrise en médecine expérimentale
offert à l'Université du Québec à Chicoutimi
en vertu d'un protocole d'entente avec l'Université Laval
pour l'obtention du grade de maître es sciences (M.Se.)
FACULTÉ DE MÉDECINE
UNIVERSITÉ LAVAL
QUÉBEC
et
DEPARTEMENT DES SCIENCES HUMAINES
UNIVERSITÉ DU QUÉBEC À CHICOUTIMI
SAGUENAY
NOVEMBRE, 2006
© Cathy Cauvier, 2006

Mise en garde/Advice
Afin de rendre accessible au plus
grand nombre le résultat des
travaux de recherche menés par ses
étudiants gradués et dans l'esprit des
règles qui régissent le dépôt et la
diffusion des mémoires et thèses
produits dans cette Institution,
l'Université du Québec à
Chicoutimi (UQAC) est fière de
rendre accessible une version
complète et gratuite de cette œuvre.
Motivated by a desire to make the
results of its graduate students'
research accessible to all, and in
accordance with the rules
governing the acceptation and
diffusion of dissertations and
theses in this Institution, the
Université du Québec à
Chicoutimi (UQAC) is proud to
make a complete version of this
work available at no cost to the
reader.
L'auteur conserve néanmoins la
propriété du droit d'auteur qui
protège ce mémoire ou cette thèse.
Ni le mémoire ou la thèse ni des
extraits substantiels de ceux-ci ne
peuvent être imprimés ou autrement
reproduits sans son autorisation.
The author retains ownership of the
copyright of this dissertation or
thesis. Neither the dissertation or
thesis, nor substantial extracts from
it, may be printed or otherwise
reproduced without the author's
permission.

RÉSUMÉ
La population de la Gaspésie est issue de fondateurs d'origines diversifiées qui ont pu
transmettre à leurs descendants des déterminants génétiques qui leur sont propres. Le but
de cette recherche est d'établir les caractéristiques des bassins génétiques des descendants
de quatre populations fondatrices de la Gaspésie (Acadiens, Canadiens-français, Anglo-
normands et Loyalistes Britanniques) à partir d'analyses généalogiques et d'analyses
moléculaires de polymorphismes neutres de l'ADN mitochondrial. Ces analyses ont été
effectuées sur un corpus de 399 participants répartis selon leur origine ethnique. Des
valeurs d'apparentement élevées ainsi que les indices de diversité moléculaires les plus
faibles chez les participants acadiens suggèrent que l'effet fondateur est plus marqué au
sein de ce groupe. En opposition, la plus grande hétérogénéité est présente chez le groupe
d'ascendance anglo-normande. Cette première étude démogénétique de la population
gaspésienne laisse donc entrevoir la présence de bassins génétiques distincts tout en
ouvrant la voie à de futures études épidémiologiques.

REMERCIEMENTS
Prendre une décision n'est que le début du voyage dont on ignore tout du point
d'arrivée... Lorsque je me suis inscrite, je n'avais qu'une vague idée de ce qui m'attendait.
Maintenant, je sais que tous ceux qui ont franchi cette étape du 2e cycle ont eu cette
réflexion que la somme des connaissances accumulées au cours de l'exécution de leur
projet de recherche était largement inférieure à celles qu'ils avaient accumulées sur eux-
mêmes. Réaliser une maîtrise, c'est un cadeau qu'on se fait à soi-même. Une aventure,
c'est le mot que j'utiliserais pour décrire ces deux années. Une aventure, parfois très
difficile, mais une aventure quand même et, d'abord et avant tout, avec soi. Pendant ce
« marathon », j'ai vécu des petits bonheurs et traversé des moments très difficiles au cours
desquels j'ai avancé en aveugle. Mais, même si on se sent parfois bien seule sur le
parcours, on ne l'est jamais et heureusement
! ! !
A toutes ces personnes qui ont prononcé la
bonne parole au bon moment : MERCI INFINIMENT
! ! !
Plus personnellement, je souhaite remercier ma directrice de maîtrise Hélène Vézina
qui,
outre le fait de m'avoir proposé le projet Patrimoine génétique des Gaspésiens et de
m'avoir incité à me dépasser et à me questionner tout au long de sa réalisation, a fait preuve
d'une grande compréhension en respectant mon besoin de latitude lorsque l'appel du large
devenait trop fort :"Merci de cette marque de confiance et de m'avoir souvent permis de
partir... pour mieux revenir. " Un merci également à mon co-directeur Damian Labuda qui
m'a accueilli le temps d'un été dans son laboratoire à Sainte-Justine : l'opportunité
d'assimiler ces techniques m'a certes confrontée à leurs difficultés, mais m'a permis aussi
d'en comprendre les principes.

IV
Merci à Ève-Marie Lavoie (pour ton regard au-delà..., ton positivisme, ta gentillesse,
ton équilibre et ton soutien moral informatique), à Diane Brassard (pour ton humanité, pour
les soirées à discuter autour d'une tasse de thé à la marocaine, pour le beau Mao...et pour
tant d'autres choses encore...) et à Lise Gobeil (dont la discrétion n'a d'égal que la
gentillesse, ce qui n'est pas peu dire
!!!!!).
Merci à Claudia Moreau (pour ta patience, ton
enthousiasme !!!) et à Vania Yotova du laboratoire de Sainte-Justine.
Merci à l'équipe de généalogistes supervisée par France Néron, à Michèle Jomphe, à
Sophie Claveau, à Jean-Philippe T. Archibald et à Bernard Casgrain pour votre
disponibilité et votre gentillesse. Merci également à Louis Houde et à Jean-Luc Allard
pour leur soutien informatique et leur patience ainsi qu'à Marc Tremblay pour sa rigueur
scientifique et sa curiosité. Merci finalement aux recruteurs et aux participants sans qui
cette étude n'aurait pas eu lieu ainsi qu'aux organismes subventionnaires qui ont supporté
ce projet (CRSNG, RMGA et FRSQ).
Un merci spécial à Julie qui a été mise sur ma route pour une raison que je commence
à peine à comprendre. Merci d'avoir été mon guide à travers ma tempête: je t'en serai
éternellement reconnaissante.
À ma famille, à Steph, à Carole, à Pascal, à Edith, à Anthony ainsi qu'à tous mes
amis électroniques : « Même à des centaines de kilomètres, savoir que vous étiez avec moi,
m'a très souvent donné la force de tenir le coup. Merci de tout cœur pour vos petits mots,
vos courriels, vos appels. »
À mon coin de pays: mon inspiration, ma destination...
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
1
/
174
100%