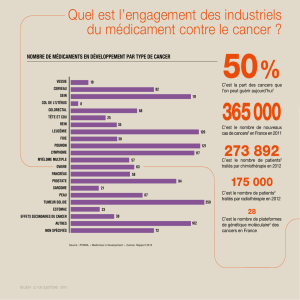
Génétique

Génétique
T. Lecomte et P. Laurent-Puig
Introduction
Les nombreux et rapides progrès réalisés dans la connaissance du génome
humain ont permis une meilleure compréhension de l’origine génétique des
cancers. Le cancer est une maladie de l’ADN qui résulte de l’accumulation d’al-
térations génétiques et, plus particulièrement, de gènes impliqués dans la
prolifération et la différenciation cellulaire. L’ensemble de ces événements a été
particulièrement bien mis en évidence dans la forme sporadique du cancer
colorectal ayant abouti au schéma maintenant classique de la cancérogenèse
multi-étape (1). La meilleure connaissance du processus de carcinogenèse a
aussi permis de déterminer des « profils génétiques tumoraux » associés au
pronostic, qui dans un avenir proche, seront susceptibles d’influencer les
modalités de la prise en charge d’un patient atteint d’un cancer digestif en
termes de traitement et de surveillance. Parallèlement, l’oncogénétique a vu sa
place grandir en cancérologie digestive grâce à la découverte de gènes de prédis-
position héréditaire aux cancers digestifs, incluant des cancers rares comme les
tumeurs endocrines du tube digestif, mais aussi les cancers les plus fréquents
comme le cancer colorectal. La génétique offre donc de nouveaux outils qui
permettent, non seulement de mieux comprendre les mécanismes des cancers
digestifs, mais aussi de dépister les sujets ayant une prédisposition génétique au
cancer, avec comme corollaire la mise en place de stratégies de prévention.
Les bases moléculaires de la cancérogenèse
du tube digestif : l’exemple du cancer colorectal
Au moins deux mécanismes différents de carcinogenèse colorectale ont été
individualisés à partir de données issues de l’étude des altérations génétiques
somatiques des cancers colorectaux (2). Le mécanisme le plus fréquent est
caractérisé par une instabilité chromosomique se traduisant par la perte

récurrente de segments chromosomiques. L’autre mécanisme est caractérisé par
une instabilité génétique liée à un défaut de réparation de l’ADN. Ces deux
mécanismes concernent la plupart des cancers colorectaux sporadiques et aussi
ceux s’intégrant dans les deux formes de prédisposition familiale au cancer
colorectal les plus fréquentes que sont la polypose adénomateuse familiale
(PAF) et le syndrome HNPCC (hereditary non polyposis colorectal cancer). Bien
que ces deux mécanismes soient différents et ciblent des gènes distincts, les
voies de signalisation impliquées lors de la transformation maligne des cellules
épithéliales coliques sont les mêmes dans les deux groupes de cancers colorec-
taux (3). Il s’agit essentiellement des voies de signalisation APC/β-caténine,
TGF-β, RAS et TP53. Cette classification nosologique est intéressante non
seulement sur le plan de la compréhension de la carcinogenèse colorectale,
mais aussi sur le plan de la prise en charge des patients atteints d’un cancer
colorectal car elle est associée au pronostic de la maladie.
Les altérations génétiques somatiques des cancers colorectaux
(tableau I)
Le phénotype d’instabilité chromosomique ou phénotype LOH+ (Loss of
heterozygosity)
Environ 85% des cancers colorectaux sporadiques présentent un phénotype
LOH+ caractérisé par des pertes alléliques sur les bras courts des chromo-
somes 8 (8p) et 17 (17p) et sur les bras longs des chromosomes 5 (5q), 18
(18q) et 22 (22q) (2,4). Des mutations ponctuelles inactivatrices des gènes
suppresseurs de tumeurs TP53 et APC, respectivement localisés sur le bras
court du chromosome 17 et sur le bras long du chromosome 5, sont fréquem-
24 Les cancers digestifs
Tableau I – Anomalies génétiques des cancers colorectaux en fonction du phénotype LOH+ et MSI+.
Tumeur LOH+ Tumeur MSI+
Hyperploïde Diploïde
Pertes alléliques sur les chromosomes 17p, 5q, Peu ou pas de pertes alléliques
18q, 8p, 22q
Mutations fréquentes des gènes TP53, APC Mutations rares des gènes APC, TP53
Mutations fréquentes de l’oncogène KRAS2 Mutations fréquentes de l’oncogène BRAF
Altérations des gènes de la réparation des
mésappariements de l’ADN (hMSH2, hMLH1,
hMSH6, hMLH3 et hMSH3)
Mutations des gènes TGFRII, BAX, TCF4,
β
-caténine
Instabilité chromosomique Instabilité génétique

ment associées à ces pertes alléliques (4). La conséquence de cette association
d’altérations au niveau d’un gène (perte allélique et mutation ponctuelle inac-
tivatrice délétère au niveau de l’allèle conservé) est une perte de la fonction de
ce gène. Dans ce type de cancer, les cellules tumorales présentent un contenu
anormal en ADN (aneuploïdie) correspondant le plus souvent à une hyper-
ploïdie consécutive à une multiplication anormale de l’ADN sans division
cellulaire (endoreduplication). Les mécanismes moléculaires à l’origine de cette
instabilité chromosomique sont en partie expliqués par les altérations de la
protéine APC qui joue un rôle dans le contrôle de la stabilité chromosomique
au cours de la division cellulaire (5). La protéine APC normale interagit à la
fois avec les microtubules en maintenant leur polymérisation, et avec la
protéine EB1 qui se fixe aux kinétochores des chromosomes. Les mutations
inactivatrices du gène APC conduisent à la synthèse d’une protéine tronquée
qui perd ses sites de liaison aux microtubules et à la protéine EB1 dont les
conséquences sont des anomalies de ségrégation des chromosomes responsables
de la perte de certains de leurs fragments. Cependant, l’inactivation du gène
APC n’est probablement pas suffisante pour provoquer l’instabilité chromoso-
mique, et d’autres gènes sont probablement impliqués, en particulier ceux
participant au contrôle du fuseau mitotique lors de la division cellulaire.
Le phénotype d’instabilité génétique ou phénotype MSI+ ou RER+
Les cancers colorectaux appartenant à ce groupe sont caractérisés par des alté-
rations génétiques liées à un défaut de réparation de l’ADN se traduisant par
une instabilité des loci microsatellites (6). Les tumeurs appartenant à ce groupe
ont un phénotype dit MSI+ (MicroSatellite Instability) ou, anciennement,
RER+ (Replicative ERror). Sur le plan anatomo-pathologique, ces cancers sont
préférentiellement localisés au niveau du côlon proximal, souvent peu diffé-
renciés, et présentent souvent une mucosécrétion abondante et un stroma riche
en lymphocytes (7). Dans ce groupe de tumeurs, les cellules tumorales ont un
contenu normal en ADN (normoploïdie ou diploïdie). Les gènes impliqués
dans ce mécanisme de carcinogenèse sont les gènes du système MMR
(MisMatch Repair) qui participent à la réparation des mésappariements
de l’ADN, essentiellement représentés par les gènes hMSH2, hMLH1 et
hMSH6 (2). Les microsatellites sont des séquences d’ADN constituées de la
répétition en tandem d’un motif de 1 à 4 nucléotides. Ils sont remarquable-
ment abondants et uniformément distribués dans l’ensemble du génome
humain, localisés en majorité dans des régions non codantes de l’ADN et, du
fait de leur structure répétée, difficiles à répliquer. Au cours de la réplication de
l’ADN, ils sont des cibles privilégiées d’erreurs de l’ADN-polymérase respon-
sables de mésappariements de l’ADN. Mais ces erreurs sont normalement
réparées par les protéines du système MMR. Dans les cancers où le système
MMR est déficient, les microsatellites deviennent instables du fait de l’accu-
mulation d’erreurs de réparation de l’ADN. La méthode de référence pour la
Génétique 25

détermination du statut MSI est un génotypage de 5 loci microsatellites (7). Il
s’agit d’un test simple de biologie moléculaire qui peut être réalisé à partir
d’ADN extrait d’un fragment tumoral fixé et inclus en paraffine, à condition
que la fixation soit réalisée dans un liquide de type aldéhyde qui n’altère pas
l’ADN contrairement au Bouin. La réalisation de ce test nécessite générale-
ment une comparaison entre l’ADN tumoral et l’ADN non tumoral, ce qui
suppose un contrôle histologique de la qualité du prélèvement tumoral. En cas
de phénotype MSI+, des altérations de taille des loci microsatellites testés
seront détectées par la méthode PCR (Polymerase Chain Reaction).
L’immunohistochimie est une méthode alternative et indirecte pour détecter
un phénotype MSI+ basée sur la mise en évidence d’une extinction des
protéines hMSH2, hMLH1 ou hMSH6 au niveau des cellules tumorales (8).
Les mécanismes moléculaires responsables de l’inactivation des gènes MMR
sont de plusieurs types : mutations ponctuelles, délétions ou hyperméthylation.
La méthylation de l’ADN est une altération dite épigénétique liée à la « sénes-
cence » de l’ADN susceptible de modifier l’expression des gènes et de conduire
à la transformation cellulaire maligne (9). La méthylation des cytosines conte-
nues dans les îlots CpG, situés au niveau du promoteur de certains gènes, est
capable d’inhiber leur expression. Une hyperméthylation acquise du promo-
teur du gène hMLH1 est fréquemment observée dans les cancers colorectaux
sporadiques MSI+ des sujets âgés (10). De nombreux gènes impliqués dans des
voies de contrôle du cycle cellulaire, de l’apoptose et de la réparation de l’ADN
sont inactivés par la survenue de mutations liées à des défauts de réparation de
mésappariements de l’ADN dans des séquences répétées codantes de quelques
paires de bases de ces gènes. Les principaux gènes cibles de ces mutations sont
le gène du récepteur de type II du TGF-β, les gènes pro-apoptiques BAX ou
CASPASE-5, les gènes des facteurs de transcription TCF-4 ou CDX2 (11). En
revanche, la fréquence des mutations des gènes TP53 et APC est significative-
ment moins fréquente que celle observée dans les cancers LOH+.
La carcinogenèse colorectale : un processus multi-étape (fig. 1)
Le cancer colorectal est un modèle de choix pour l’étude de la chronologie des
événements moléculaires impliqués dans les différentes étapes de la transfor-
mation maligne d’une cellule épithéliale colique à travers l’étude des foyers de
cryptes aberrantes, des adénomes et des cancers de différents stades (1). Les
étapes de la transformation maligne ont été particulièrement bien étudiées dans
les cancers colorectaux LOH+. Concernant les cancers MSI+, les différentes
étapes de la carcinogenèse sont moins bien connues. À noter que la répartition
sur le cadre colique des différents « phénotypes » tumoraux n’est pas homo-
gène. En effet, les cancers MSI+ sont beaucoup plus fréquents au niveau du
côlon proximal qu’au niveau du côlon distal, et les cancers du côlon distal sont
à plus de 95% des cancers LOH+.
26 Les cancers digestifs

Valeurs pronostiques des altérations moléculaires du cancer
colorectal
L’établissement du pronostic, chez les patients traités pour un cancer colorectal,
repose actuellement sur quelques éléments simples anatomo-pathologiques,
mais ceux-ci sont imparfaits. Les données actuelles sur la caractérisation molé-
culaire des cancers colorectaux laissent entrevoir la perspective de nouveaux
paramètres pronostiques parmi les altérations génétiques des cancers colorec-
taux (12). Outre l’intérêt pronostique de cette approche moléculaire dans la
prise en charge du cancer colorectal, ce type d’approche pourrait permettre de
prédire l’effet des traitements.
Les pertes alléliques 8p, 17p et 18q ont été rapportées dans plusieurs études
comme étant associées de façon indépendante à un pronostic péjoratif (12).
Celles des bras 8p et 18q semblent être les plus discriminantes en termes de
survie globale et sans récidive des cancers colorectaux de stades II-III (13-15).
La valeur pronostique des pertes alléliques n’est pas seulement qualitative.
L’importance quantitative des pertes alléliques a été rapportée comme étant un
facteur pronostique de survie péjoratif des cancers colorectaux de stades II et
III, ce qui rejoint la valeur pronostique péjorative de l’aneuploïdie (16). Les
mutations somatiques du gène suppresseur de tumeur TP53, de l’oncogène
KRAS et l’hyperméthylation du promoteur du gène suppresseur de tumeur p16
ont également une valeur pronostique péjorative (17-19). Le résultat le plus
clairement démontré concerne la valeur pronostique favorable du statut MSI+.
Génétique 27
Fig. 1 –
Les deux voies principales de la carcinogenèse colorectale.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
1
/
21
100%