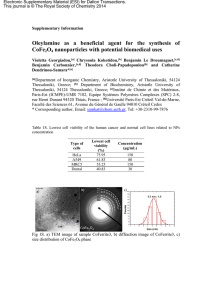
et al - divine [id]

Comment réconcilier les aspects moléculaires
cellulaires et tissulaires du cancer ?
Jean-Pascal Capp
LISBP, UMR CNRS 5504, UMR INRA 792
INSA/Université de Toulouse
France
capp@insa-toulouse.fr 22ème séminaire de l’AMOC, Nice, 22/05/2016

Cell, 1990
Maintenant :
Yates & Campbell,
Nat Rev Genet, 2012
Science, 1976
Les origines du modèle génétique actuellement dominant

Stratton, 2011, Science
Les objectifs du séquençage massif

Garraway & Lander, 2013, Cell
De grands projets de séquençage (2010-2015)

Alexandrov et al, 2013, Nature
Et un nombre incroyablement élevé de mutations détectées
Forte hétérogénéité inter-tumorale, en nombre et en type de mutations
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
1
/
58
100%