iv le complement

I. DEFINITION
II. LES BARRIÈRES ÉPITHÉLIALES
III. LES RÉCEPTEURS
IV. LE COMPLÉMENT
V. LES CELLULES
VI. LES AC NATURELS
VII. L'INFLAMMATION
VIII. IMMUNITÉ INNÉE ET DÉFENSE ANTI-INFECTIEUSE
IX. ROLE DE L'IMMUNITE INNEE DANS LES REPONSES ADAPTATIVES
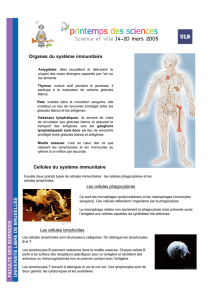
L'IMMUNITE NATURELLE OU INNEE

L'immunité innée ou naturelle comprend dans sa définition la
plus simple les mécanismes de défense intrinsèque contre les
infections microbiennes.
•Première apparue dans l'évolution
•Non spécifique de l'agent infectieux, reconnaît des structures
communes à des micro-organismes non présents chez
l'homme. Pour une même structure, discrimine entre germes
extérieurs et commensaux.
•Préexiste à la première introduction du pathogène, ses
éléments sont présents chez l'individu sain
•Immédiatement efficace, première ligne de défense.
•Interagit avec l'immunité spécifique qu'elle aide à recruter.
I. DEFINITION

I. COMPOSANTS DE
L'IMMUNITE INNEE
Cellules épithéliales
Récepteurs cellulaires
Activation cellulaire
Phagocytose
Récepteurs humoraux
Réaction inflammatoire
Ac naturels
Lymphocytes NK, NKT, T-γδ

II. BARRIERES EPITHELIALES

II. BARRIERES
EPITHELIALES,
les défensines
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
1
/
72
100%