Chapitre 4 : La cellule bactérienne, paroi et structures

Chapitre 4 : La cellule
bactérienne, paroi et
structures
I. La paroi cellulaire
Les bactéries sont majoritairement divisées en deux groupes selon leur type de paroi,
les Gram+ et les Gram-. Le type de paroi d’une espèce peut en effet être défini par
l’observation des bactéries au microscope après une coloration de Gram (Fig.2), qui colore
les cytoplasmes des bactéries Gram+ en violet et des Gram- en rose. La différence de couleur
obtenue par la coloration de Gram s’explique par une différence structurelle au niveau de la
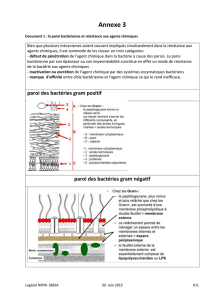
paroi des bactéries (Fig.1). Les bactéries Gram+ ont une paroi très épaisse, et une seule
membrane. Les bactéries Gram- ont une paroi globalement plus fine et plus fragile, mais
deux membranes lipidiques. Cette différence a une origine évolutive, puisqu’en effet ces
deux groupes sont éloignés au niveau évolutif ; l’identification du type de membrane d’une
espèce bactérienne permet donc de la classer dans l’un des deux grands groupes évolutifs
des bactéries.
Dans les deux cas, le peptidoglycane est un composant essentiel de la paroi
bactérienne. Ce complexe formé de sucres et de protéines forme un maillage régulier et
solide qui participe à la solidité de la paroi et donc à la résistance de la bactérie aux
changements environnementaux. Chez les Gram+, le peptidoglycane est épais (20 à 80
nanomètres) et situé à l’extérieur de l’unique membrane plasmique. Chez les Gram-, il est
plus fin (5 à 8 nanomètres) et situé entre les deux membranes interne et externe.
La membrane plasmique, qui est donc unique chez les Gram+ et double chez les
Gram-, est composée d’une bicouche lipidique, tout comme la membrane des eucaryotes.
L’espace située entre les deux membranes des bactéries Gram- est appelé le périplasme ;
leurs deux membranes sont appelées membranes interne et externe. Des protéines sont
ancrées sur les deux membranes des Gram- et celle des Gram+. Ces protéines peuvent avoir

des parties exposées vers le cytoplasme, le périplasme ou le milieu extérieur. Elles servent à
la communication entre les bactéries, la détection des conditions environnementales, la
prise de nutriments dans le milieu, le déplacement, ou encore la sécrétion de protéines vers
l’extérieur.
Les bactéries Gram- possèdent une grande molécule externe appelée
lipopolysaccharide (LPS). Celle-ci est composée d’une partie lipidique qui permet son
ancrage dans la membrane externe et d’une longue chaîne de sucres. Cette molécule
participe à la stabilisation de la paroi des Gram-. De façon intéressante, sa composition est
très différente entre les espèces et légèrement différentes entre les différentes souches
d’une même espèce. Quand deux souches d’une même espèce ont un LPS différent, on parle
de sérotypes différents. Le LPS est donc souvent utilisé comme marqueur d’identification
d’un échantillon bactérien. De plus, cette molécule est facilement reconnue par le système
immunitaire humain, et sa détection entraîne une inflammation et une réponse
immunitaire ; le LPS est, du point de vue humain, un élément important de la réponse
immunitaire face aux infections bactériennes.

Fig. 1 : Structure de la paroi des bactéries à Gram négatif et à Gram positif.
II.
Fig. 2 : Principe de la coloration de Gram.
L’étape dans l’alcool permet la décoloration des cellules, uniquement si elles sont Gram-,
l’alcool ne pouvant franchir la couche de PDG des Gram+. La contre-coloration à la safranine
marque les deux types de cellules mais ne se voit que sur les Gram- parce qu’elles ont été
Coloration au
cristal violet
Décoloration
à l’alcool 95°
Contre-
coloration à
la safranine
Violet : Gram+
Rose : Gram-
Milieu
extracel
lulaire
Membr
ane
externe
Péri
plas
me
Cy
top
las
me
Memb
rane
intern
e
Milieu
extracel
lulaire
Membr
ane
plasmiq
ue
Cy
top
las
me
Protéines
Phospholipides
(membrane)
Peptidoglycane
LPS
Légende :
Gram négatives
Gram positives

décolorées ; le rose ne se voit pas sur le fond déjà violet des Gram+. La couleur des cellules
est déterminée par observation au microscope.
II. Les structures de surface
Plusieurs types de structures peuvent être observables en surface chez certaines
espèces sous certaines conditions seulement. On parle de structures de surface
optionnelles. Les gènes correspondants peuvent ne pas être présents chez toutes les
espèces d’un même genre, voire même toutes les souches d’une même espèce. Ces gènes
peuvent aussi n’être exprimés que sous certaines conditions, comme par exemple le
manque ou l’abondance de nutriments dans le milieu, la présence d’un support d’adhésion,
ou encore les conditions d’humidité.
Le flagelle est une structure de taille importante, sa longueur pouvant atteindre le
double de la longueur de la cellule. Le flagelle sert à la motilité (avec un T !), c’est-à-dire la
capacité de la bactérie à nager en milieu liquide. Les bactéries peuvent avoir un ou plusieurs
flagelles. Leur disposition peut être différente selon les espèces : monotriche (à une seule
extrémité de la cellule), amphitriche (aux deux pôles de la cellule), péritriche (tout autour de
la cellule) ou lophotriche (en touffe à une extrémité). Les bactéries sans flagelles sont parfois
appelées atriches. Au sein d’une seule espèce, toutes les bactéries présentent la même
disposition flagellaire. Le flagelle peut être observé au microscope optique après coloration à
la fuschine de Leifson.

Fig. 3 : Représentation des différentes morphologies flagellaires possibles.
Le filament du flagelle est constitué d’une protéine appelée flagelline. La base du
flagelle assure son ancrage dans la paroi, à travers les membranes plasmiques et le
peptidoglycane. Le flagelle tourne sur lui-même à une vitesse importante (200 à 1000
tours/min). Cette propulsion est assurée par l’énergie tirée de la force protomotrice, c’est-à-
dire l’énergie créée par la sortie des électrons issus du métabolisme, hors de la cellule. La
vitesse qui peut être atteinte par les bactéries flagellées est extrêmement importante,
certaines espèces se déplaçant à 60 fois leur longueur par seconde. Par comparaison, Usain
Bolt atteint 6,2 fois sa taille par seconde, dans un milieu (l’air) beaucoup moins visqueux que
l’eau ; un guépard en pleine course peut atteindre 25 fois sa longueur par seconde.
Les pili (singulier : pilus) sont des structures de surface composés de protéines
assemblées en filaments, les pilines. On distingue d’abord les pili communs ou fimbriae,
courts, qui servent à l’attachement aux surfaces et aux déplacements en milieux semi-solide
(motilité par « swarming » ou « gliding »). Les pili communs recouvrent toute la surface de la
bactérie.
Les pili sexuels sont beaucoup plus longs, bien que composés aussi de pilines ; il n’y en a
généralement qu’un par cellule. Ils sont encodés par des gènes présents sur certains
plasmides, les plasmides conjugatifs (mécanisme de conjugaison). Les pilis sexuels sont
produits par les bactéries porteuses de ces plasmides (bactérie donneuse), et servent à
Monotriche Lophotriche Amphitriche Péritriche
 6
7
8
9
10
6
7
8
9
10
1
/
10
100%