D - Stud`Ant STAPS

INTRODUCTION
Niveaux d’organisation structurale
1) Niveau chimique
- Le plus simple
- Les atomes se combinent pour former des molécules.
1) Niveau cellulaire
- Cellules = unités structurales et fonctionnelles de base
- Très grande variabilité dans la taille et la forme des cellules => diversité des fonctions
des cellules.
1) Niveau tissulaire
- Les organismes les plus simples ne sont constitués que d’une seule cellule mais chez
d’autres organismes, la structure se complexifie jusqu’au niveau tissulaire.
- Les tissus sont des groupes de cellules semblables ayant la même fonction.
- Chaque type de tissu joue un rôle particulier dans l’organisme => diversité.
1) Niveau organique
Ce niveau organique résulte de la jonction de différents types de tissus. Les organes sont
des structures composées d’au moins deux tissus différents et dotés de fonctions définies
et de forme reconnaissable stable.
1) Niveau des systèmes
- Système ou appareil composé d’un ensemble d’organes travaillant ensemble pour une
même fonction.
1) Niveau de l’organisme
Le niveau de l’organisme représente l’ensemble de tous les niveaux de complexité
travaillant en synergie pour assurer le maintien de la vie.

Composition chimique de l’organisme
I- Les atomes
Ils sont les plus petites unités de matière qui composent toute l’existence chimique. Une
substance pure qui ne contient qu’un seul type d’atome est un élément chimique. Chaque élément
chimique est représenté par un symbole comportant une ou deux lettres. Parmi la centaine
d’éléments chimiques naturels, seulement 24 sont essentiels à la structure et au fonctionnement de
notre organisme.
L’hydrogène, l’oxygène, le carbone et l’azote sont les principaux éléments puisqu’ils représentent
plus de 99 % des atomes du corps humain.
Ensuite, nous trouvons 7 éléments minéraux qui sont les substances les plus abondantes dissoutes
dans les liquides de l’organisme. Enfin, il y a les oligo-éléments (13) qui se trouvent en infime
quantité dans l’organisme (0,01 %) mais qui sont néanmoins essentiels à la croissance et au
fonctionnement normal de l’organisme.
Les propriétés chimiques de ces atomes dépendent de 3 particules qui les constituent ; Les
protons, les neutrons et les électrons.
Les protons et les neutrons sont localisés dans les noyaux des atomes et les électrons gravitent
autour du noyau.
I- Les molécules
Deux ou plusieurs atomes liés les uns aux autres forment une molécule. La plupart des molécules
de l’organisme contiennent du carbone (ce sont les molécules organiques). Lorsqu’une molécule
ne contient pas d’atome de carbone, on l’appelle molécule inorganique. La plupart des molécules
organiques de notre organisme se classent en quatre groupes :
- Les glucides.
- Les lipides.
- Les protides.
- Les acides nucléiques.
A) Les glucides

Ils représentent 1 % de la masse corporelle totale. Mais ils jouent un rôle capital dans les
réactions chimiques qui fournissent de l’énergie aux cellules. Ils sont composés de carbone,
d’hydrogène et d’oxygène.
1) 1) Les monosaccharides : les oses :
Les glucides les plus simples sont les monosaccharides.
Formule générale d’un monosaccharide : (CH2O) n
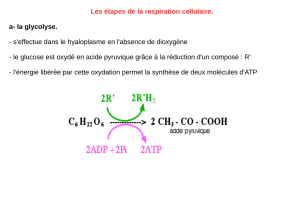
Ex : le glucose avec 6 atomes de carbones : C6H12O6.
Ce n’est qu’une formule brute. Le glucose peut également être présenté sous d’autres formes.
Lorsqu’il contient 6 atomes de carbone, on dit que c’est un hexose.
1) 2) Les disaccharides : les osides
: La liaison de deux monosaccharides constitue un disaccharide (ex : le saccharose).
1) 3) Les polysaccharides : les polyosides :
Lorsqu’un grand nombre de monosaccharides se lient les uns aux autres, il y a formation de
polysaccharide. Dans toutes les cellules vivantes, le glucose est stocké sous forme de
polysaccharides. Dans les cellules végétales, ce polysaccharide s’appelle l’amidon et dans les
cellules animales, le glycogène (hydrolyse qui permet la libération d’unité de glucose).
A) Les lipides
Ils sont des composés qui représentent 15 % de la masse corporelle totale. Ils sont constitués
d’atomes de carbone et d’hydrogène. Les lipides peuvent être subdivisés en quatre sous-classes.
1) Les acides gras :
Un acide gras est constitué d’une chaîne d’atomes de carbone portant à l’une de ses extrémités un
radical carboxyle (-COOH). Lorsque les atomes de carbone de la chaîne carbonée sont reliés par
des liaisons simples, l’acide gras est un acide gras saturé.
Formule générale d’un acide gras saturé : CH3-(CH2) n-COOH avec 2<n<22
Certains acides gras contiennent une ou plusieurs double-liaisons dans leur chaîne carbonée. Ce
sont donc des acides gras insaturés.
Formule générale d’un acide gras insaturé : CH3-(CH2) n1-CH=CH-(CH2) n2-COOH
Lorsqu’un acide gras ne contient qu’une seule double-liaison, c’est un acide gras mono insaturé.
S’il y en a plusieurs, c’est un acide gras polyinsaturé.
1) Les triglycérides :
Ils contiennent la majeure partie des lipides de l’organisme. Ils sont constitués par la liaison de
trois acides gras qui se lient à une molécule de glycérol. Ces trois acides gras ne sont pas

forcément identiques. Ainsi, une grande variété de triglycérides peut être constituée.
1) Les phospholipides :
Ils ont une structure générale semblable à celle des triglycérides mais ils comportent une
différence importante. Dans le phospholipide, le glycérol est associé à deux acides gras et un
groupement phosphate.
1) Les stéroïdes :
Ils ont une structure nettement différente des autres sous-groupes de lipides (ex : le cholestérol).
A) Les protides (protéines)
Ils représentent 17 % de la masse corporelle totale. Ils sont constitués d’atomes de carbone,
d’hydrogène, d’oxygène et d’azote. Les protéines sont constituées par des acides aminés qui se
lient entre eux par l’intermédiaire de liaisons peptidiques.
➔ Acides aminés :
Formule générale d’un acide aminé :
=> H2N : groupement aminé.
=> O=C-OH : groupement acide carboxylique.
=> R : Radical, chaîne latérale.
Il existe 20 acides aminés différents ayant chacun une chaîne latérale différente. Le corps humain
peut tous les synthétiser sauf neuf d’entre eux qui sont des acides aminés dit essentiels et qui
doivent donc être apportés par l’alimentation.
➔ Les polypeptides :
Les acides aminés, constituant les protéines vont s’assembler en polymères c’est-à-dire en
chaînes d’acides aminés. Les chaînes d’acides aminés sont formées par la liaison de radical
carboxyle d’un acide aminé au radical aminé d’un autre acide aminé.
L’union des deux acides aminés donne un dipeptide. L’union de trois acides aminés donne un
tripeptide et l’union de dix ou plusieurs acides aminés donnent un polypeptide.
Les molécules qui contiennent plus de 50 acides aminés => protéines. On dit donc que les
protéines sont des macromolécules. La succession de ces acides aminés constituent une chaîne
polypeptidique qui constitue primaire.
A) Les acides nucléiques : ADN et ARN
Ces molécules sont importantes car elles sont responsables de la transmission de l'information

génétique. Il y a deux classes d'acides nucléiques :
L'ADN : acide désoxyribonucléique.
L'ARN : acide ribonucléique.
Ces deux types d'acides nucléiques sont constitués de séquences polymères qui se répètent.
Chaque unité est appelé un nucléotide. Un nucléotide comporte trois composants : un groupement
phosphate, un glucide et une base azotée.
1) ADN :
Les nucléotides de l'ADN contiennent un pentose (= un sucre) qui est le désoxyribose. On trouve
dans l'ADN 4 nucléotides différents qui correspondent aux 4 bases azotées qui peuvent se lier au
désoxyribose. Ces 4 bases se subdivisent en 2 classes :
Les bases puriques : Adénine ou Guanine.
Les bases pyrimidiques : Cytosine ou Thymine.
Les deux chaînes de nucléotides sont enroulées en double hélice. Ces deux chaînes sont reliées
entre elles par des liaisons hydrogène entre les bases. Les bases ne s’unissent pas au hasard :
A se lie à T.
C se lie à G.
On dit que A et T sont des bases complémentaires tout comme C et G.
1) ARN :
La structure de l'ARN est différente de celle de l'ADN en quelques points seulement :
La molécule d'ARN n'est composée que d'une seule chaîne.
Le désoxyribose est remplacé par le ribose.
La Thymine est remplacée par l’Uracile.
A) L’adénosine triphosphate : ATP.
C'est une énergie utilisable par toutes les formes d'organisme. Toutes les cellules doivent
fabriquer elles-mêmes leurs molécules d'ATP. La molécule d'ATP est un nucléotide de l'ARN
contenant de l'Adénine plus deux groupements phosphate. Les groupements phosphates en
excédant sont rattachés par des liaisons chimiques uniques appelées liaisons phosphates. Ces
liaisons ont la particularité d'être très riche en énergie. C’est la rupture de ces liaisons par
hydrolyse qui va permettre de libérer de l'énergie qui peut être utilisée immédiatement pour
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
1
/
42
100%