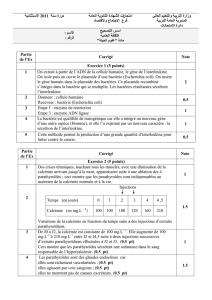
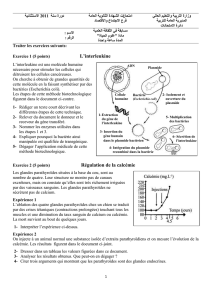
Exemple 3 - Gold Mastering

(a)
\
0 10
{b)
Système sensoriel somatique 425
Figure
à
(b)
al. ,

432 Systèmes sensoriel et moteur
Système moteur somatique
Figure

Cerveau et comportement : aspects neurochimiques 51 3
LES VO l ES DE DÉCOUVERTE
L'éveil de ma passion pour la dopamine ...
1
1'
À
Figure A à

À
� . �
��
. .
. � .
0 0
�
�
789

805
 6
6
1
/
6
100%