Aménagement forestier durable

138
LA SOCIÉTÉ PROVANCHER D’HISTOIRE NATURELLE DU CANADA
LA DIVERSITÉ FLORISTIQUE
Hélène D’Avignon est ingénieur forestier.
Introduction
Depuis le Sommet de Rio en 1992, les nations recon-
naissent l’importance d’agir devant les menaces d’épuise-
ment mondial des ressources forestières. Elles se demandent
comment continuer à les exploiter sans pour autant nuire
à leur capacité naturelle de renouvellement. C’est à partir
de cette préoccupation commune qu’est né, partout dans le
monde, le concept d’aménagement forestier durable (AFD).
Au Canada comme au Québec, le concept de durabilité
repose sur six principes, ou critères, qui sont évalués au
moyen d’indicateurs déterminés. Par exemple, pour s’as-
surer de conserver la diversité biologique (critère 1) du
territoire forestier québécois, le ministère des Ressources
naturelles propose de recourir à huit indicateurs (MRN,
1996).
L’industrie forestière est de plus en plus incitée à
adhérer à ces principes de l’AFD. La mesure incitative la plus
en vue est la certification des produits du bois par « éco-label-
lisation ». L’éco-labellisation garantit qu’un produit provient
d’une forêt aménagée selon les principes de l’AFD. Les indus-
triels sont de plus en plus nombreux à vouloir s’engager dans
un processus de certification, malgré sa complexité.
Un des défis de taille auquel une industrie forestière
doit faire face en choisissant la voie de l’AFD est de conser-
ver la diversité biologique dans son aire commune, tout en
poursuivant ses activités de récolte. En effet, ces dernières
affectent vraisemblablement la diversité à deux niveaux de
l’écosystème forestier : celui du paysage forestier, lors de la
planification des récoltes, et celui du peuplement lui-même,
lors de la récolte proprement dite. On doit donc, dans ces
territoires, choisir des indicateurs qui permettent de mesurer
la diversité à ces deux niveaux. Les objectifs de conservation
doivent de plus être en accord avec les objectifs de conserva-
tion aux niveaux supérieurs de l’organisation biologique,
comme le paysage régional (Saucier et al., 1998). On ne sau-
rait donc prétendre conserver la diversité biologique sans
tenir compte de la hiérarchie des écosystèmes au sein des-
quels elle s’organise.
Si beaucoup d’énergie a été déployée jusqu’à ce jour
pour élaborer les critères et les indicateurs à l’échelle régio-
nale, l’expérimentation de mesures, au Québec du moins,
est à peine amorcée dans les grands territoires destinés à la
récolte (aires communes). C’est dans ce contexte que nous
avons voulu mettre sur pied une méthode rigoureuse de
mesures et de suivi d’indicateurs de diversité biologique dans
une aire commune située en forêt boréale. Ainsi, les indus-
triels pourront s’appuyer sur l’image de la diversité révélée
par les premières mesures des indicateurs pour établir leurs
objectifs de conservation, et des mesures subséquentes pour
vérifier s’ils les ont atteints.
Nous présentons les indicateurs, essentiellement des
constituants de l’écosystème. Nous décrivons ensuite les
méthodes de mesure employées et l’image que les indica-
teurs livrent de la diversité végétale du territoire. Enfin, nous
voyons en quoi cette image peut aider à l’établissement d’ob-
jectifs de conservation de la diversité biologique. Cet article
présente une synthèse d’un projet d’élaboration de métho-
des de mesure et de suivi d’indicateurs d’AFD qui a été
expérimenté dans l’aire commune 031-07 entre 1997 et 2000
(D’Avignon et al., 1999, 2000 ; Ouimet et al., 2000 ; Périé
et al., 2000). Les résultats rapportés ici proviennent de ce
projet. Mais avant, il faut dire quelques mots sur ce territoire
boréal.
Territoire
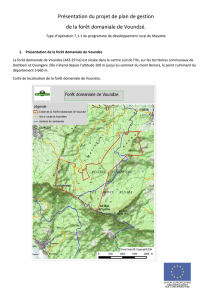
L’aire commune 031-07 est située dans la réserve
faunique des Laurentides au nord de la ville de Québec
(figure 1). La végétation y est entièrement boréale : sapins,
épinettes et bouleaux blancs forment massivement le pay-
sage forestier, duquel pointent ça et là quelques peuplements
de mélèzes et de trembles. Depuis toujours, la mosaïque des
peuplements dans ce paysage doit son agencement et sa com-
position aux attaques marquées de la tordeuse des bourgeons
de l’épinette et, dans une moindre mesure, aux feux, dont
le plus important remonte à 1878. Ce n’est que récemment
que l’exploitation forestière est venue ajouter régularité et
discontinuité dans le modèle jadis irrégulier de cette mosaï-
que de peuplements. Le relief est par ailleurs très accidenté ;
plus du quart du territoire est caractérisé par des pentes qui
dépassent 30 % de déclivité. L’altitude moyenne est d’environ
800 m dans la partie sud du territoire et autour de 900 m dans
Aménagement forestier durable :
ÉVALUATION D’INDICATEURS DE LA BIODIVERSITÉ DANS
UNE AIRE COMMUNE
Hélène D’Avignon

139
LE NATURALISTE CANADIEN, VOL. 125 No 3 AUTOMNE 2001
LA DIVERSITÉ FLORISTIQUE
Figure 1. Localisation de l’aire commune 031-07, de la zone d’étude et des points d’observation de l’inventaire écologique
du MRN. Les points sur la figure représentent le centre des virées composées de cinq à sept points d’observation.
sa partie nord. Les glaciers ont modelé le relief en collines
moutonnées, en vallées et en cuvettes. Ils y ont déposé du till
en grande majorité (92 %) et, par leurs eaux de fonte, un peu
de dépôt fluvio-glaciaire. C’est la présence des montagnes
qui fait que le climat est plus rude qu’il ne l’est habituellement
à cette latitude. La température moyenne annuelle n’est que
de – 0,6 °C et les précipitations sont abondantes : 1 500 mm
dont 600 mm sous forme de neige (Anonyme, 1997). Les
sols portent la marque de toutes ces influences : climat froid
et humide, relief très accidenté et végétation boréale. Ils sont
classés en podzols ferro-humiques et humo-ferriques selon
l’altitude, et plusieurs d’entre eux sont caractérisés par des
horizons indurés (Lamontagne et Nolin, 1997), comme ceux
que l’on trouve plus au nord.
Indicateurs et méthodes d’évaluation
Un des objectifs inhérents au projet était de choisir
des indicateurs pour lesquels nous puissions disposer de
données. Du ministère des Ressources naturelles, nous avons
utilisé les cartes écoforestières ainsi que les données d’inven-
taire écologique de terrain prises au cours de l’été 1989. Nous
avons également récupéré la carte écologique et la typologie
écologique de la végétation d’un projet de gestion intégrée
des ressources, réalisé dans ce territoire en 1994 (MEF, 1994 ;
tableau 1).
Au cours des dernières années, la diversité de deux
des constituants majeurs de l’écosystème forestier a vrai-
semblablement été touchée dans ce territoire : la diversité
de la mosaïque des peuplements, si on se place à l’échelle du
paysage forestier, et la diversité des sous-bois, si on se place à
l’échelle du peuplement.
Échelle du paysage forestier
À l’échelle du paysage, le premier indicateur consis-
tait donc en l’évaluation de la proportion de chaque peu-
plement au sein de la mosaïque du point de vue de 1) sa
composition en essences ; 2) sa composition en essences par
âge et 3) sa composition en essences par âge et par classe de
drainage (tableau 2). Nous en avons produit les cartes en
compilant les données cartographiques numérisées dont

140
LA SOCIÉTÉ PROVANCHER D’HISTOIRE NATURELLE DU CANADA
LA DIVERSITÉ FLORISTIQUE
Échelle de la mosaïque
2. forme des polygones
Échelle du peuplement
indice de forme (IF) du polygone p/r à une
surface de référence (cercle)
Composition végétale Groupement
d'essences/classe d'âge 18 catégories (p. ex. SBB 50)
Configuration des
groupements
d'essences/classe d'âge
1. classe de taille
des peuplements fréquence des polygones de ≤ 10 ha jusqu'à ≥ 100 ha
Fragmentation 1. indice d'isolement distance moyenne (m) entre deux polygones du
même groupement d'essences/classe d'âge
2. indice de distribution
spatiale distribution aléatoire, régulière ou groupée
Diversité des écosystèmes 1. richesse typologique 32 types forestiers (p.ex. pessière noire à mousses)
2. diversité des
types forestiers indice de similarité d'espèces entre les types forestiers
Diversité en
espèces vasculaires richesse spécifique
végétale indice du nombre maximal d'espèces vasculaires à
l'intérieur d'un type forestier
Diversité des gènes des
espèces vasculaires rareté locale pourcentage d'espèces vasculaires observées
une seule fois dans les inventaires
Indicateur Indice Description
Tableau 2. Indicateurs mesurés à l’échelle de la mosaïque et à l’échelle du peuplement dans l’aire commune 031-07
Tableau 1. Données utilisées lors du projet d’expérimentation de méthodes de mesures et de suivi d’indicateurs de la
diversité végétale dans l’aire commune 031-07
Sources des données Description
Échelle de la mosaïque
Carte écoforestière Cartes forestières numérisées (1:20 000) du MRN de 1990, contenant
l’information suivante : groupements d’essences par classe d’âge, par classe de
hauteur et par classe de densité.
Carte écologique Carte numérisée (1:50 000) du projet de Gestion intégrée des ressources
(MEF, 1994) contenant l’information suivante : étage bioclimatique, district
écologique, dépôt meuble, drainage et modificateur de drainage.
Échelle du peuplement
Inventaire écologique du MRN Base de données phytosociologiques de 388 points d’observation provenant de
65 virées de l’inventaire écoforestier, réalisé entre le 24 juillet et le 20 septembre
1989 dans les régions écologiques 8f et 9c de Thibault (1987). Recensement des
espèces selon une liste préétablie.
Inventaire du projet de Gestion Base de données phytosociologiques de 512 points d’observation distribués
intégrée des ressources (MEF, 1994) dans l’aire commune par groupes de cinq à sept, sur des transects le long des
caténas. Recensement des espèces les plus abondantes ou reconnues pour avoir
une signification écologique.

141
LE NATURALISTE CANADIEN, VOL. 125 No 3 AUTOMNE 2001
LA DIVERSITÉ FLORISTIQUE
MINISTÈRE
DES
RESSOURCES
NATURELLES
Sapinière à mousses et à oxalis
nous disposions. Le deuxième indicateur consistait en la
caractérisation de la mosaïque elle-même, essentiellement
sa configuration et son degré de fragmentation. Pour ce faire,
nous avons calculé cinq indices en utilisant comme données
le périmètre et la surface des polygones cartographiques des
peuplements. Trois indices de configuration ont permis de
caractériser respectivement la taille, la forme et l’agencement
de la mosaïque des peuplements, tandis qu’un indice d’iso-
lement et un de connexité ont permis d’évaluer son degré de
fragmentation.
Afin d’évaluer les niveaux de modification de la
mosaïque des peuplements forestiers depuis l’intensifica-
tion des activités humaines, nous l’avons comparée avec la
mosaïque qui caractérisait le paysage forestier avant cette
intensification. Pour ce faire, nous nous sommes servis de
la description de la forêt primitive qu’ont fait Leblanc et
Bélanger (1997) dans un secteur du territoire de l’aire com-
mune.
Échelle du peuplement
À l’échelle du peuplement, nous avons d’abord dressé
un portrait de la diversité floristique de chacun en compi-
lant les espèces végétales qui avaient été recensées dans les
inventaires écologiques de terrain (tableau 2). Cependant,
ce portrait général ne pouvait constituer en soi un indicateur
de diversité satisfaisant. Nous étions plutôt intéressés à con-
naître d’abord la variété des écosystèmes qui se trouvaient
dans le territoire, puis de caractériser certains aspects de
leur diversité de composition en espèces vasculaires. C’est
pourquoi nous avons fait de la typologie écologique l’objet
du premier indicateur de diversité
à cette échelle. La typologie résulte,
en effet, du classement des associa-
tions régulières d’espèces observées
(sous-bois) sur le terrain, en fonc-
tion des conditions environnantes
qui y prévalent. Il s’agit donc d’une
typologie d’écosystèmes (composée
de types appelés couramment types
forestiers), puisque l’idée d’écosys-
tème s’impose dès qu’on constate
que des espèces coexistent et intera-
gissent avec leur environnement.
Sous un peuplement de sapi-
nière apparemment homogène, par
exemple, différents sous-bois, libel-
lés comme suit, peuvent s’y trou-
ver : « oxalis », « mousses-oxalis »,
« mousses », « mousses-sphai-
gnes », « sphaignes ». La typologie
ne renseignait cependant que sur
un aspect de la diversité, celui qui
réfère à la richesse, c’est-à-dire à la
quantité de types forestiers propres
à ce territoire. Nous savions donc combien il y en avait,
mais nous ignorions où et dans quelle proportion ils s’y
trouvaient. La réalisation d’une carte était une façon de le
savoir, l’estimer à tout le moins. Mais comment identifier
des sous-bois à partir d’une photo aérienne ? Comme nous
ne pouvions pas les identifier par ce moyen classique de
cartographie, nous les avons déduits à partir des relations
qu’ils ont avec les éléments du milieu environnant qui sont
en mesure d’être photo-interprétés : la hauteur, l’âge et la
nature des arbres, le dépôt meuble et la classe de drainage.
Pour cartographier les sous-bois, nous avons procédé indi-
rectement, mais objectivement, en utilisant un modèle sta-
tistique de prédiction (D’Avignon et al., en préparation). Le
modèle permettait de déterminer un sous-bois particulier
avec une précision connue à partir des données de terrain
énumérées ci-dessus.
Nous avons ensuite caractérisé la diversité de com-
position en espèces vasculaires des types forestiers à l’aide
de trois indices qui se rapportent chacun grossièrement aux
trois niveaux d’expression de la diversité biologique :
1. indice de diversité d’écosystèmes : échelle de similarité de
composition en espèces entre les types forestiers ;
2. indice de diversité d’espèces vasculaires : estimation de la
richesse maximale en espèces vasculaires de chaque type
forestier ;
3. indice de diversité de gènes (isolement des populations
d’espèces) : pourcentage d’espèces observées une seule fois
dans les inventaires de terrain.
Le portrait de la diversité livré par ces trois indices a
constitué le deuxième indicateur de diversité à l’échelle du

142
LA SOCIÉTÉ PROVANCHER D’HISTOIRE NATURELLE DU CANADA
LA DIVERSITÉ FLORISTIQUE
Échelle du paysage
MINISTÈRE
DES
RESSOURCES
NATURELLES
peuplement. Ces indices avaient l’avantage de ne nécessiter
dans les calculs que des données de recensement d’espèces.
Ce choix de mesures de diversité s’inspire du principe selon
lequel la présence des plantes vasculaires ou des bryophytes
est le meilleur indicateur des écosystèmes terrestres (Pielou,
1993). L’énoncé de ce principe contient l’idée que les espèces
végétales reflètent non seulement la diversité biologique
mais la créent. De par leur nature et leur nombre, elles engen-
drent une diversité de niches ou d’habitats, depuis le feuillage
dans les arbres jusqu’au tapis des mousses au sol, que vien-
nent coloniser quantité d’autres espèces, tant animales que
végétales. Cette diversité de niches ou d’habitats résulte elle-
même des conditions du milieu (climat, géologie, topogra-
phie), qui viennent, dans une plus large vue, déterminer sa
répartition spatiale.
Le premier indice a été obtenu en soumettant les
données de végétation à une analyse dite de groupements
(D’Avignon et al., 2000). Le résultat se présente sous la forme
d’une échelle synthétique de similarité d’espèces sur laquelle
les types forestiers vont des plus aux moins ressemblants.
Le deuxième indice se rapporte à la richesse maximale
de chacun des types forestiers, seule richesse qui puisse faire
l’objet de comparaison entre eux. Nous ne pouvions, en effet,
comparer valablement les richesses entre elles en prenant le
nombre d’espèces observées dans les placettes de terrain parce
que celui-ci est corrélé directement avec la quantité de placet-
tes. En effet, la théorie suppose que plus la surface du territoire
d’un type d’écosystème donné est grande, plus on y recense
d’espèces. Cependant, après une certaine superficie couverte,
la probabilité de trouver de nouvelles espèces diminue pro-
gressivement, jusqu’à devenir à peu près nulle. En illustrant
graphiquement ce taux de diminu-
tion de nouvelles espèces trouvées en
fonction de la superficie de territoire
couverte, on obtient une courbe de
type asymptotique, de laquelle on peut
dériver une équation. L’asymptote est
ce point sur la courbe qui représente le
nombre maximal d’espèces qu’il serait
à peu près possible de trouver dans un
type d’écosystème donné dans les limi-
tes d’un territoire donné. Nous avons
donc déterminé les équations de chaque
type forestier à l’aide des espèces recen-
sées dans leurs placettes pour obtenir
cette richesse maximale représentée par
l’asymptote. Un test statistique a permis
de vérifier que la richesse maximale ne
dépendait pas du nombre de placet-
tes grâce auquel elle avait été estimée.
C’est ainsi que nous avons pu comparer
objectivement la richesse entre les types
forestiers.
Le pourcentage d’espèces recensées une seule fois
dans les inventaires apporte une certaine mesure de l’iso-
lement des populations d’individus de ces espèces dans le
territoire. C’est en ce sens que cet indice peut se rapporter à
la diversité des gènes. En effet, le risque d’appauvrissement
génétique est susceptible d’augmenter si une population
d’une de ces espèces, constituée de peu d’individus, deve-
nait si éloignée d’une petite population de la même espèce
qu’aucun échange de pollen ne puisse se produire. Cepen-
dant, comme les inventaires d’espèces desquels découle le
calcul de cet indice n’étaient pas exhaustifs, il nous a fallu
l’interpréter avec discernement.
Portrait de la diversité végétale
Comment tirer la meilleure interprétation possible
du portrait de la diversité végétale du territoire par le calcul
de tous ces indices ? En commençant par examiner les carac-
téristiques de la mosaïque des peuplements, ensuite celles du
peuplement, soit en procédant du général au particulier.
Échelle du paysage forestier
Composition de la mosaïque de peuplements
En 1990, le territoire de l’aire commune contient la
gamme des peuplements forestiers les plus fréquemment
observés dans cette partie de la forêt boréale : bétulaies pures
et mélangées, sapinières de couverts variés, pessières, mélé-
zaies et aulnaies. L’ensemble des sapinières occupe un peu
plus de 60 % de la superficie – 34 % d’entre elles étant des
sapinières pures –, et les peuplements dits en régénération
 6
7
6
7
1
/
7
100%