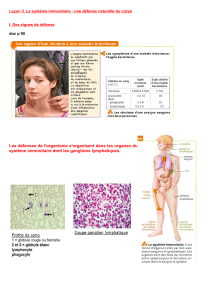
I. Les lymphocytes T

T
TH
HÈ
ÈS
SE
E
En vue de l'obtention du
D
DO
OC
CT
TO
OR
RA
AT
T
D
DE
E
L
L’
’U
UN
NI
IV
VE
ER
RS
SI
IT
TÉ
É
D
DE
E
T
TO
OU
UL
LO
OU
US
SE
E
Délivré par l'Université Toulouse III - Paul Sabatier
Discipline ou spécialité :
Immunologie
JURY
Pr. Denis Hudrisier
Dr. Emmanuel Donnadieu
Dr. Jacques Bertoglio
Dr. Loïc Dupré
Ecole doctorale : Biologie, Santé, Biotechnologies (BSB)
Unité de recherche : Dynamique moléculaire des interactions lymphocytaires
Directeur(s) de Thèse : Dr. Loïc Dupré
Rapporteurs : Dr. Emmanuel Donnadieu
Dr. Jacques Bertoglio
Présentée et soutenue par
Fanny Lafouresse
Le jeudi 20 septembre 2012
Titre :
Implication de la protéine du syndrome de Wiskott-Aldrich
dans la migration et la rencontre des lymphocytes T CD4+
avec les cellules présentatrices de l'antigène

Sommaire
1
SOMMAIRE
REMERCIEMENTS ...................................................Erreur ! Signet non défini.
SOMMAIRE ........................................................................................................ 1
LISTE DES ABBRÉVIATIONS ........................................................................ 3
LISTE DES FIGURES ....................................................................................... 6
RÉSUMÉ .............................................................................................................. 7
ABSTRACT ......................................................................................................... 8
AVANT-PROPOS ............................................................................................... 9
INTRODUCTION .............................................................................................11
I. Les lymphocytes T : chefs d’orchestre de l’immunité adaptative, une vie
bien rythmée ......................................................................................................12
1. Développement et sélection thymique des lymphocytes T dans les organes
lymphoïdes primaires ........................................................................................................... 14
a) Expression du pré-TCR à la membrane des thymocytes .......................................... 15
b) La sélection positive ................................................................................................. 16
c) La sélection négative ................................................................................................ 17
d) Tolérance périphérique par les lymphocytes T régulateurs ..................................... 17
e) Lymphocytes T CD8+ et lymphocytes T CD4+ ........................................................ 18
f) Sortie du thymus et recirculation ............................................................................. 18
2. Extravasation des lymphocytes T dans les organes lymphoïdes secondaires .......... 19
a) « Roulement » des lymphocytes T induit par une adhésion aux HEV de faible
intensité .................................................................................................................... 21
b) Activation des récepteurs aux chimiokines .............................................................. 22
c) Adhésion des lymphocytes T aux HEV induite par l’engagement de LFA-1 .......... 25
d) Passage des HEV...................................................................................................... 27
e) Contrôle de l’entrée lymphocytaire .......................................................................... 30
3. Rencontre avec les cellules présentatrices de l’antigène dans les organes
lymphoïdes secondaires ........................................................................................................ 31
a) Organisation des organes lymphoïdes secondaires .................................................. 32
b) Caractérisation de la mobilité des lymphocytes T dans les organes lymphoïdes
secondaires ............................................................................................................... 33
c) Caractérisation de la rencontre avec les cellules dendritiques dans les organes
lymphoïdes secondaires ........................................................................................... 39
d) Rencontre avec les cellules dendritiques en condition de non-infection ................. 40
e) Rencontre avec les cellules dendritiques en condition d’infection .......................... 46
4. Activation des lymphocytes T CD4+ ....................................................................... 50
a) Architecture des synapses immunologiques ............................................................ 51
b) Signalisation lymphocytaire ..................................................................................... 58
c) Terminaison de l’interaction cellulaire .................................................................... 59
5. Phase effectrice des lymphocytes T CD4+ ............................................................... 59
a) Devenir des lymphocytes T CD4+ dicté par les cellules présentatrices d’antigène . 59
b) Les lymphocytes T CD4+ effecteurs dans les organes lymphoïdes secondaires ...... 60

Sommaire
c) Les lymphocytes T CD4+ effecteurs hors des organes lymphoïdes secondaires ..... 61
II. La protéine du syndrome de Wiskott-Aldrich : un régulateur clef du
cytosquelette d’actine des lymphocytes T .......................................................62
1. Le cytosquelette d’actine ......................................................................................... 63
a) Polymérisation et dépolymérisation des filaments d’actine ..................................... 63
b) Molécules et modèles pour l’étude du cytosquelette d’actine .................................. 64
c) Implication du cytosquelette d’actine dans les fonctions des lymphocytes T .......... 65
d) Organisation du cytosquelette d’actine lors de la migration amiboïde .................... 65
e) Organisation du cytosquelette d’actine lors de la rencontre avec les cellules
présentatrices de l’antigène ...................................................................................... 69
f) Organisation du cytosquelette d’actine lors de la formation de la synapse
immunologique ........................................................................................................ 70
g) Les protéines régulatrices du cytosquelette d’actine ................................................ 71
2. La protéine du syndrome de Wiskott-Aldrich (WASP) .......................................... 71
a) Structure et partenaires protéiques de WASP .......................................................... 71
b) Expression et localisation de WASP dans les cellules hématopoïétiques ................ 74
3. Le syndrome de Wiskott-Aldrich (WAS) ................................................................ 75
a) Historique de la découverte de WAS ....................................................................... 76
b) Pathologies ............................................................................................................... 76
c) Manifestations cliniques ........................................................................................... 79
d) Défauts cellulaires .................................................................................................... 82
e) Thérapie .................................................................................................................... 85
OBJECTIFS DU TRAVAIL ............................................................................91
RÉSULTATS ..................................................................................................... 92
DISCUSSIONS GÉNÉRALES ET PERSPECTIVES ................................. 132
I. Rôle de WASP dans la dynamique des lymphocytes T : point de vue
fondamental .....................................................................................................133
1. Au niveau cellulaire ............................................................................................... 133
2. Au niveau moléculaire ........................................................................................... 142
II. Rôle de WASP dans la dynamique des lymphocytes T : point de vue
physiopathologie..............................................................................................147
ANNEXES ........................................................................................................ 151
BIBLIOGRAPHIE .......................................................................................... 183

LISTE DES ABBREVIATIONS
ADP Adenosin DiPhosphate
AIRE AutoImmune Regulator
APECED Autoimmune PolyEndocrinopathy Candidiasis Ectodermal Dystrophy
Arp2/3 Actin-related protein 2/3
ATP Adenosine TriPhosphate
Ca2+ Calcium
CALDAG Calcium and diacyglycerol-regulated
CD- Cluster de Différenciation-
CGD Chronic granulomatous disease
CMH Complexe Majeur d’Histocompatibilité
CRIB Cdc42-Rac Interactive Binding
CTS Central Terminaison Sequence
DAG 1,2 Diacylglycerol
DARC Duffy Antigen Receptor for Chemokines
DC-SIGN Dendritic Cell-Specific Intercellular adhesion molecule-3-Grabbing
Non-integrin
EBV Epstein Barr Virus
ERM Ezrine, Radixine, Moésine
ESAM Endothelial Cell Selective Adhesion Molecule
EVH1 Ena-VASP-Homology-1 domain
FDC Follicular Dendritic Cells
FRC Fibroblastic Reticular Cells
GDP Guanosine 5’ DiPhophate
GEF Guanine nucleotide Exchange Factor
GTP Guanosine 5’ Triphosphate
HDMVEC Human Dermal Microvascular-Endothelial Cells
HEC High Endothelial Cells
HEV High Endothelial Venules
HLA Human Leukocyte Antigen
HLMVEC Human Lung Microvascular- Endothelial Cells
HUVEC Human Umbilical Vein-Endothelial Cells
ICAM-1 InterCellular Adhesion Molecule-1

4
IL- InterLeukine-
JAM- Junctional Adhesion Molecule-
LAD Leucocyte Adhesion Deficiency
LBRC Lateral Border Recycling Compartement
LFA-1 Lymphocytes Function-associated Antigen-1
LTR Long Terminal Repeat
MAdCAM-1 Mucosal Addressin Cellular Adhesion Molecule-1
Min Minute
MLCK Myosin Light-Chain Kinase
MMLV Moloney Murine Leukemia Virus
MPSV MyeloProliferative Sarcoma Virus
MTOC MicroTubule Organizing Center
N-WASP Neural-Wiskott-Aldrich Syndrome Protein
PBS Primer Binding Site
PBSF Pre-B-cell growth Stimulating Factor
PECAM-1 Platelet/Endothelial Cell Adhesion Molecule-1
PIP5K PhosphatidylInositol-4-Phosphate 5 Kinase
PLC Phospholipase C
plt Paucity in Lymph-node T cells
PNAD Peripheral Node ADdressin
PPT PolyPurine Tract
PRD Prolin Rich Domain
PTX Pertussis ToXine
RAG Recombinaison Activating Genes
SCID Severe Conbined Immunodefiency Diseases
SDF-1 Stromal cell-derived factor 1
SMAC Supra Molecular Activation Cluster
S1P1 Sphingosine 1 Phosphate 1
TCR T Cell Receptor
Th T Helper cells
TIRF Total Internal Reflexion Fluorescence
TSLP Thymic Stromal LymphoPoïetine
VAP1 Vascular Apoptosis-Inducing Protein 1
VCAM Vascular Cell Adhesion Molecule
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
1
/
215
100%